
Carcharhinus mancinae, Ebersole & Cicimurri & Stringer, 2019
|
publication ID |
https://doi.org/ 10.5852/ejt.2019.585 |
|
publication LSID |
lsid:zoobank.org:pub:181B6FBA-ED75-4BB4-84C4-FB512B794749 |
|
DOI |
https://doi.org/10.5281/zenodo.3664579 |
|
persistent identifier |
https://treatment.plazi.org/id/C940D627-E666-474A-8383-B6B9F5A4D01E |
|
taxon LSID |
lsid:zoobank.org:act:C940D627-E666-474A-8383-B6B9F5A4D01E |
|
treatment provided by |
Plazi |
|
scientific name |
Carcharhinus mancinae |
| status |
sp. nov. |
Carcharhinus mancinae sp. nov.
urn:lsid:zoobank.org:act:C940D627-E666-474A-8383-B6B9F5A4D01E
Galeocerdo alabamensis – Thurmond & Jones 1981: 63 , fig. 28, left.
Galeocerdo latidens – Manning 2003: 374 , fig. 21.3.2.
Etymology
The species name is dedicated to the late Lois Nickey Mancin of Birmingham, Alabama, USA, in honor of her years of volunteer service to MSC and her life-long dedication to science education.
Material examined
Holotype
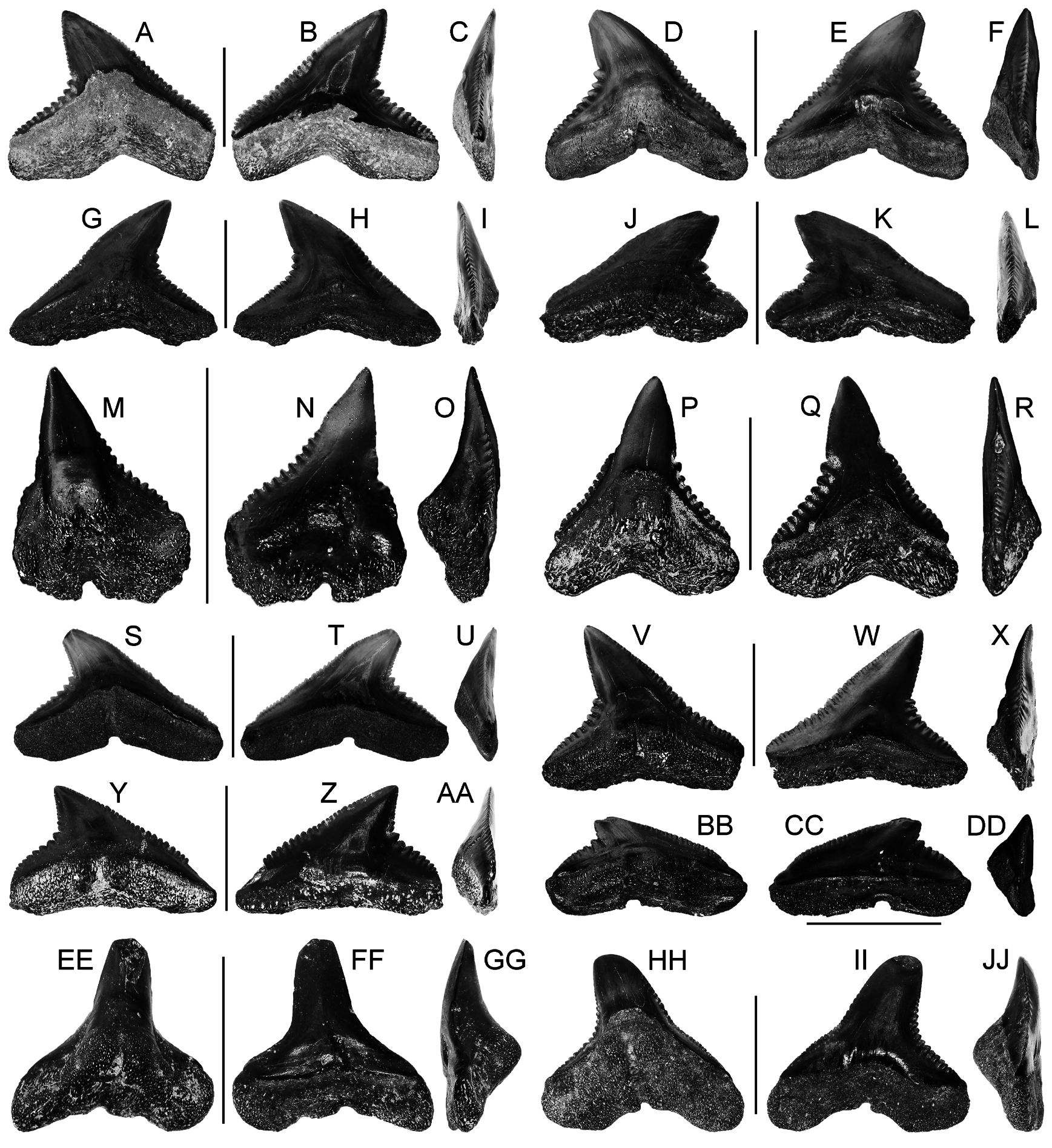
UNITED STATES OF AMERICA – Alabama • upper right lateral tooth; Claiborne Group ; MSC 39036. ( Fig. 25 View Fig A–F).
Paratypes
UNITED STATES OF AMERICA – Alabama • 1 upper left anterior tooth; same collection data as for holotype; MSC 39037 ( Fig. 25 View Fig G–K) • 1 upper left posterior tooth; same collection data as for holotype; MSC 39038 ( Fig. 25 View Fig V–Z) • 1 lower right anterior tooth; same collection data as for holotype; MSC 39040 ( Fig. 25 View Fig L–P) • 1 lower left lateral tooth; same collection data as for holotype; MSC 39041 ( Fig. 25 View Fig Q–U) .
Referred material
UNITED STATES OF AMERICA – Alabama • 98 isolated teeth; Claiborne Group ; ALMNH PV1993.3.408, ALMNH PV 1996.1.9, ALMNH PV2016.3.74, MSC 188.15 , MSC 188.27 , MSC 188.40 , MSC 188.89 , MSC 12653, MSC 12728, MSC 36952.1 – 3 , MSC 37448, MSC 37502, MSC 37522.1 – 3 , MSC 37532, MSC 37896, MSC 38416, MSC 38423, MSC 38424.1 – 9 , MSC 38425.1 – 2 , MSC 38426.2 – 4 , MSC 38426.6 – 20 , MSC 38426.22 – 58 , MSC 38503.1 – 5 , MSC 38546, MSC 38558.1 – 2 , MSC 38798, MSC 38863 (6 specimens) .
Stratum typicum
Basal Gosport Sand, middle Bartonian, Zone NP17.
Locus typicus
Site ACl-4, Clarke County, Alabama, USA.
Differential diagnosis
This species can be differentiated from most other Claiborne Group chondrichthyans by the presence of compound and/or double serrations, which are absent on all taxa except Galeocerdo clarkensis White, 1956 and Otodus (Carcharocles) sp. Further separated from the carcharhiniforms Abdounia Cappetta, 1980 , Galeorhinus , Hemipristis , Negaprion , Pachygaleus , and Physogaleus by the presence of fine mesial and distal serrations that extend to the apex of the main cusp. Differentiated from Galeocerdo clarkensis by the presence of erect and triangular upper anterior teeth, lower lateral teeth with a narrower and more erect crown, uniformly concave distal cutting edge with compound serrations medially, and lateral teeth have a more pronounced and ‘pinched’ lingual root protuberance. This species is separated from the following Paleogene Carcharhinus sensu stricto Underwood & Gunter (2012) species: the lower Lutetian Carcharhinus marçaisi from northern Africa ( Arambourg 1952) by having serrated cutting edges; the middle Eocene Carcharhinus sp. from Jamaica ( Underwood & Gunter 2012), the middle-to-late Eocene Carcharhinus sp. from Egypt ( Underwood et al. 2011), and the middle-to-late Eocene Carcharhinus underwoodi and Carcharhinus sp. from Madagascar ( Samonds et al. 2019) by having compound and/or double serrations and a more ‘pinched’ and pronounced lingual root boss; the Priabonian Carcharhinus balochensis from Pakistan Adnet et al. 2007 by having a less concave distal cutting edge, shorter and mesiodistally thinner crown in all anterior and lateral tooth positions, and pointed (as opposed to rounded) serrations on the lower two-thirds of the crown; and the Chattian Carcharhinus perseus Adnet et al., 2007 by having compound serrations and mesiodistally narrower crown in all tooth positions.
Description
UPPER ANTERIOR TEETH. Teeth with broad, triangular crown. Crown erect, slightly inclined distally in anterolateral files. Mesial cutting edge elongate, slightly concave medially, slightly convex apically. Distal cutting edge elongate, concave to slightly angular medially, upper half weakly convex but less so than mesial edge. Labial crown face weakly concave medially; lingual face strongly and evenly convex. Base of the labial crown face is slightly bulbous. Crown enameloid smooth. Mesial and distal serrations coarse, weakly compound (by compound we mean one or more additional serrae located on the primary serration) and/or doubly serrate (where a serration is partially divided), extending along entire length of cutting edges. Largest mesial and distal serrations located medially on the cutting edges; serrations decrease in size both apically and basally. Root bilobate, with widely diverging, angular lobes that do not extend past mesial and distal ends of crown; distal root lobe mesiodistally wider than mesial root lobe. Root significantly higher lingually than labially. V-shaped lingual dental band. Lingual root protuberance is not well demarcated, but with wide, shallow nutritive groove.
LOWER ANTERIOR TEETH. Teeth with tall, mesiodistally narrow, triangular main cusp. Crown erect; apex with slight labial bend; cusp distally inclined in anterolateral files. Mesial and distal cutting edges elongate, sinuous, concave medially but convex basally and apically. Labial crown face is flat; lingual face is strongly convex. Mesial and distal cutting edges coarsely serrated along entire length, compound and/or doubly serrate; largest serrations located medially on both edges, reducing in size apically and basally. Root lobes short, rounded, strongly divergent, labiolingually flattened; distal root lobe slightly larger than mesial lobe. Interlobe area shallow, U-shaped. Root higher lingually than labially; lobes extend slightly beyond mesial and distal crown margins. Conspicuous but low lingual root protuberance bisected by deep nutritive groove.
UPPER AND LOWER LATERAL TEETH. Teeth with tall, triangular crown; with distally inclined cusp. Labial crown face flat, lingual face convex; both crown faces smooth. Mesial cutting edge elongate, slightly concave near the base but apical half convex. Distal cutting edge elongate, with straight apical half, strongly concave medially, transitioning to straight oblique heel. Medial concavity uniformly curving, not angular; distal heel not distinctively separated from apical half of cutting edge. Cutting edges complete across entire edge, coarsely serrated; serrations compound and/or doubly serrate. Mesially, largest serrations are located on the basal half of cutting edge, becoming finer apically. On distal edge, largest serrations located medially, slightly decreasing in size basally, significantly finer on apical half. First large serration on distal edge always compound. Root lobes short, rounded, strongly divergent. Root higher lingually than labially, with lobes extending slightly past the lateral extent of the crown. Interlobe area shallow and U-shaped. Pronounced lingual root boss bisected by deep nutritive groove.
Lower lateral teeth are similar to those in upper lateral files but differ by the following: main cusp mesiodistally narrower and shorter; mesial edge straighter; upper half of the distal edge of main cusp is more convex; and interlobe area is shallow and V-shaped rather than U-shaped.
UPPER POSTERIOR TEETH. Teeth mesiodistally wider than tall, with short, distally inclined cusp. Mesial cutting edge elongate, uniformly convex, serrated along entire length. Basal two-thirds of the distal cutting edge is weakly convex; apical one-third weakly convex. Mesial serrations generally coarsest medially; much finer basally and apically. Distal cutting edge very short, straight to weakly convex, finely serrated. Distinct distal heel forms notch with distal cutting edge; heel very coarsely serrated; serrations decrease in size distally. Serrations on cutting edges compound and/or doubly serrate. Labial crown face flat to slightly convex; lingual face is more strongly convex; both crown faces lack ornamentation. Root lobes very short, rounded, highly diverging. Very shallow U-shaped interlobe area. Weak lingual protuberance bisected by a deep nutritive groove.
Remarks
The fossil record of Carcharhinus (and Carcharhinus -like taxa) was reviewed by Adnet et al. (2007), Cappetta (2012), and Underwood & Gunter (2012). Although the origins of Carcharhinus sensu stricto Underwood & Gunter (2012) have not been resolved, the oldest representative of the genus appears to be the early Lutetian Carcharhinus marçaisi ( Arambourg, 1952) . However, the syntypes for this taxon ( Arambourg 1952: pl. 26, figs 31–48) appear better aligned with our definition of Negaprion (see below) as they are unserrated, have a narrow main cusp, and have an asymmetrical T-shaped outline. In contrast, the mancinae morphology described herein is appropriately assigned to Carcharhinus sensu stricto because it has upper teeth with a broader main cusp than those in the corresponding lower files, and all cutting edges are completely serrated to the apex. When compared to extant Carcharhinus jaw sets at MSC, SC, and figured in Voigt & Weber (2011), the teeth in our sample appear most similar to those of Carcharhinus amboinensis (Müller & Henle, 1839) and Carcharhinus leucas (Müller & Henle, 1839) , the two members of the leucas - amboinensis species group as defined by Garrick (1982). These two extant taxa have similar dentitions that are characterized by having broadly triangular upper teeth, narrower crowned triangular lower teeth that are more erect, and completely serrated cutting edges. The similarities between the C. mancinae sp. nov. teeth in our sample and the extant C. amboinensis and C. leucas led us to place our fossil teeth into the generalized tooth groups described above.
Within our sample we identified upper and lower anterior and lateral teeth, and upper posterior teeth. These tooth groups indicate that the dentition of Carcharhinus mancinae sp. nov. exhibits monognathic and dignathic heterodonty similar to the dentitions of several extant Carcharhinus species. Despite the variations in tooth morphology we observed in the fossil species, all the teeth can be attributed to the same taxon due to the presence of coarse compound and/or double serrations both mesially and distally, by having a tall and triangular main cusp with finer serrations that extend to the apex, and by the lack of a distinct distal notch on all but posterior positions. The combination of these characteristics separates these teeth from those of all other taxa in our Claiborne Group sample. The one taxon in our sample that could be confused with the new species is Galeocerdo clarkensis . However, this species lacks the dignathic heterodonty observed in Carcharhinus mancinae sp. nov., as the upper and lower teeth of G. clarkensis are difficult to differentiate, and the dentition lacks the erect triangular anterior tooth morphologies attributed to the new species. Although the upper posterior teeth of C. mancinae sp. nov. are morphologically similar to those of G. clarkensis , they can be differentiated by the absence of a distinct distal notch, and by having a distinctive ‘pinched’ lingual root protuberance that is more pronounced than that on G. clarkensis .
Adnet et al. (2007), Cappetta (2012), Underwood & Gunter (2012), and Samonds et al. (2019) recognized several species of Paleogene Carcharhinus sensu stricto. Of these, C. mancinae sp. nov. can be differentiated from the early Lutetian Carcharhinus marçaisi ( Arambourg, 1952) by having a wider crown and completely serrated cutting edges. The presence of compound and/or double serrations differentiates C. mancinae sp. nov. from the Carcharhinus sp. teeth figured by Underwood et al. (2011) from the late Eocene of Egypt and by Underwood & Gunter (2012) from the middle Eocene of Jamaica, both of which have simple serrations. The teeth of C. mancinae sp. nov. also differ from those of the Oligocene (Chattian) Carcharhinus perseus Adnet et al., 2007 by having compound and/or double serrations and a narrower crown in all tooth positions.
Carcharhinus mancinae sp. nov. closely resembles the Priabonian C. balochensis Adnet et al., 2007 from Pakistan. However, the upper lateral teeth of C. balochensis differ from those of C. mancinae sp. nov. by having a less concave distal cutting edge. Furthermore, the anterior and lateral tooth positions known for C. balochensis have a taller and mesiodistally thicker main cusp that is not as basally constricted at the base. Finally, the large serrations on the teeth of C. mancinae sp. nov. are more irregular and pointed, and the largest distal serration (located just below the finer apical serrations) projects further distally than that on C. balochensis .
Samonds et al. (2019) recently named Carcharhinus underwoodi based on teeth from middle-to-upper Eocene deposits in Madagascar. As part of their type description, Samonds et al. (2019) referred all but one of the syntypes of C. balochensis of Adnet et al. (2007) to Galeocerdo eaglesomei , the lone exception being an upper lateral tooth (fig. 3, 10–11) that the authors suggested corresponded closely to that of C. underwoodi . However, according to their type description of C. balochensis, Adnet et al. (2007) clearly stated that the lower half of the teeth have distinct double serrations, which contrasts with the simple serrations on all the teeth of C. underwoodi (D.J. Ward, pers. com.). Also, per Samonds et al. (2019), the serrations are simple in form on all middle Eocene G. eaglesomei teeth. Although the overall shape of many of the tooth positions within the dentition of C. mancinae sp. nov. appear to be shared with C. underwoodi and G. eaglesomei (see Samonds et al. 2019: figs 2–3), the compound and/or double serrations on the teeth of C. mancinae sp. nov. clearly separates them from these other taxa. In addition, the distinct ‘pinched’ morphology of the lingual root protuberance on many of the C. mancinae sp. nov. teeth also distinguish them from C. underwoodi and G. eaglesomei . Within the same assemblage, Samonds et al. (2019: figs 3s–t) also figured a tooth they referred to Carcharhinus sp. that differed from C. underwoodi by having a distinct distal notch and evenly spaced mesial serrae. The teeth of C. mancinae sp. nov. can be differentiated from this unspeciated taxon by the absence of a distal notch and by the presence of compound and/or double serrations. Finally, the triangular upper anterior tooth morphology of C. mancinae sp. nov. ( Fig. 25 View Fig G–K) is unique and appears absent in the dentitions of G. eaglesomei and all the previously described Paleogene Carcharhinus sensu stricto species.
Thurmond & Jones (1981: fig. 28, left) illustrated a tooth they referred to Galeocerdo alabamensis , but it appears this tooth belongs to C. mancinae sp. nov. Manning (2003: fig. 21.3.2) illustrated a tooth from site ACl- 4 in Clarke County that he referred to Galeocerdo latidens , but this specimen appears to be a lower lateral tooth of C. mancinae sp. nov. Citing a personal communication from David Ward, Underwood & Gunter (2012) noted a coarsely serrated species of Carcharhinus from the same locality. Photographs of these specimens provided to us by Ward confirmed they also belong to C. mancinae sp. nov.
Stratigraphic and geographic range in Alabama
The material in our sample were collected from the “upper” Lisbon Formation at site ACl-3, the basal Gosport Sand at sites ACl-4 and AMo-4, and the Gosport Sand at site ACl-15. Lower-to-middle Bartonian, zones NP16 and NP17.

| ALMNH |
Alabama Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Carcharhinus mancinae
| Ebersole, Jun A., Cicimurri, David J. & Stringer, Gary L. 2019 |
Galeocerdo latidens – Manning 2003: 374
| Manning E. M. 2003: 374 |
Galeocerdo alabamensis – Thurmond & Jones 1981: 63
| Thurmond J. T. & Jones D. E. 1981: 63 |