
Caparaonia itaiquara, Rodrigues & Cassimiro & Pavan & Curcio & Verdade & Pellegrino, 2009
|
publication ID |
https://doi.org/ 10.1206/622.1 |
|
persistent identifier |
https://treatment.plazi.org/id/C57687C1-FFE2-FFE3-B7D0-FC45FB7DFE37 |
|
treatment provided by |
Carolina |
|
scientific name |
Caparaonia itaiquara |
| status |
sp. nov. |
Caparaonia itaiquara , new species
Figures 2–5 View Fig View Fig View Fig View Fig
HOLOTYPE: MZUSP 96093 View Materials , an adult male from (20 ° 289S, 41 ° 499W): Parque Nacional do Caparaó : states of Minas Gerais and Espírito Santo: Brazil, collected by J. Cassimiro on 2 January 2005, field number MTR 10919.
PARATYPES: MZUSP 95053–65 View Materials , 97619 View Materials , from 24–30 October 2004 ; MZUSP 96088– 92 View Materials , 97620 View Materials , from 24 December 2004 to 5 January 2005; MZUSP 95635 View Materials , 98050–58 View Materials , from November 2005 ; all from Parque Nacional do Caparaó : states of Minas Gerais and Espírito Santo: Brazil, collected by J. Cassimiro and M . T. Rodrigues .
ETYMOLOGY: The specific epithet is from the Tupi language, meaning ‘‘the one that inhabits rocks’’ in reference to the habitat where this lizard is most frequently found.
DIAGNOSIS: A gymnophthalmid with ear opening and eyelid, and a slender pentadactyl limb lacking the claw on first toe. Frontonasal single; prefrontals, frontal, frontoparietals, parietals, and interparietals present. Parietals longer than wide. Collar fold absent. Three pairs of chin shields; three supraoculars, first smaller. Dorsal scales in 36–40 rows, anteriorly smooth, isodiametric or subrectangular in occipital region, becoming progressively narrower, more elongate and rounded at the arm level, and more posteriad lanceolate, strongly keeled, with sides almost juxtaposed. Occipitals present. Ventrals longer than wide, smooth, in six regular longitudinal and 21–27 transverse rows, identical in size. Scales around body 36–39; 10–14 and 15–20 infradigital lamellae under finger IV and toe IV, respectively. Males with a series of 9–10 pores without gap between preanal and femoral; femoral pores absent in females.
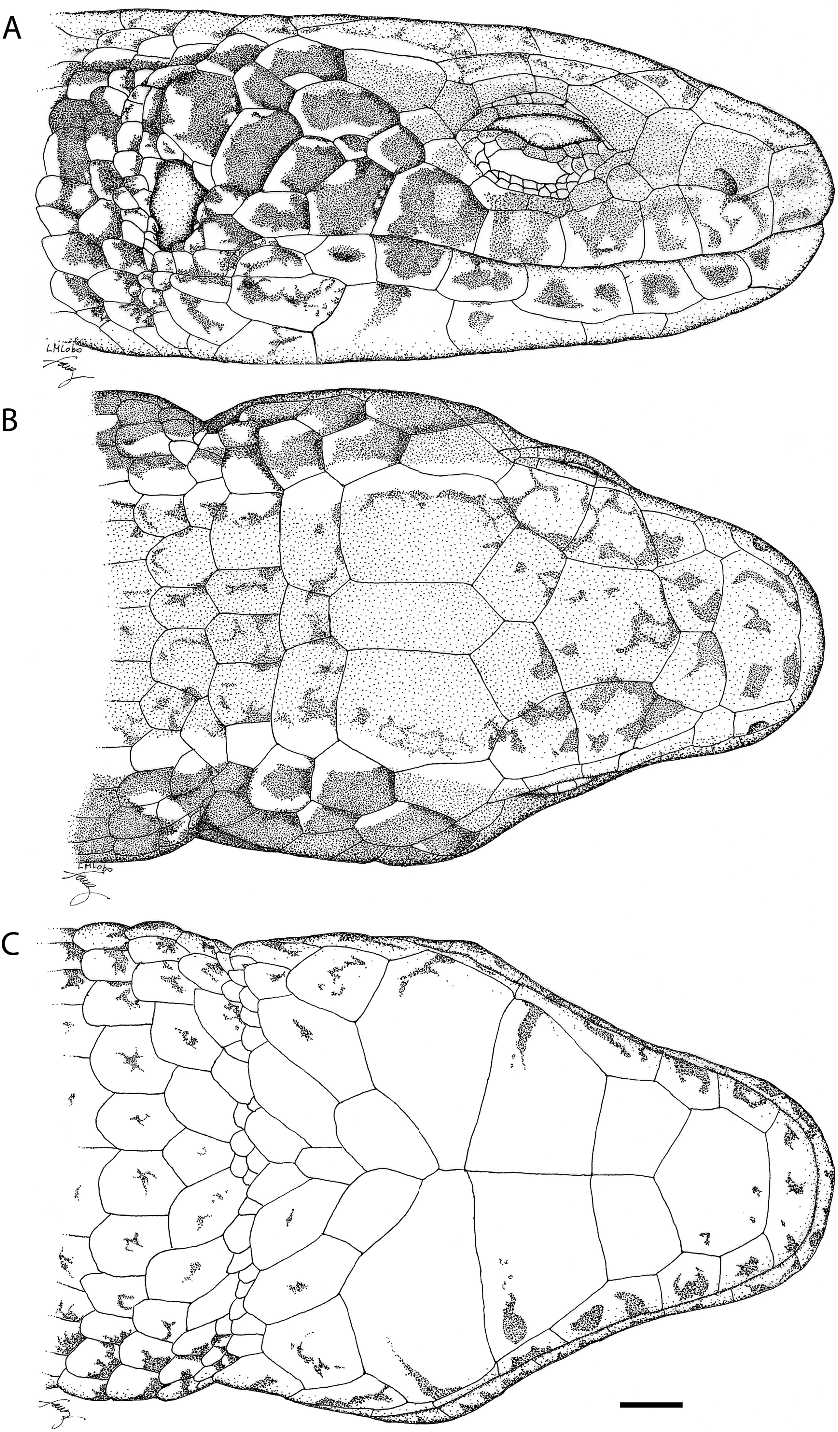
DESCRIPTION OF THE HOLOTYPE (fig. 2): Rostral scale broad, wider than high, contacting first supralabial, nasal, and frontonasal. Frontonasal pentagonal, almost twice as wide as long, contacting rostral scale, nasals, loreals, and prefrontals. Prefrontals slightly wider than long, in broad contact at midline. Frontal hexagonal, with slightly divergent lateral margins, longer than wide, slightly wider posteriorly; anteriorly indenting the prefrontal and posteriorly the frontoparietal sutures. Frontoparietals pentagonal, slightly larger than prefrontals, in slight medial contact, strongly indented by the interparietal, and in contact with second and third supraoculars. Interparietal longer than wide, as long as but much narrower than frontal, as long as but narrower than parietals. Parietals hexagonal, edged laterally by two enlarged temporal scales with approximately the same size as each other, anteriorly by the third supraocular and frontoparietal, medially by the interparietal, and posteriorly by an enlarged occipital. Posterior margin of parietals almost straight, that of interparietal rounded and extending slightly posteriorly to the parietals. A pair of distinctively enlarged occipitals separated at midline by one scale. A second pair of smaller scales following the first pair of occipitals. Three supraoculars, first the smallest, second the largest, with its largest suture with frontal, third slightly longer than second, in broad contact with frontoparietal. Nasal above first supralabial, large, slightly longer than high, with the nostril in the center and lower part of the scale, indenting suture with labial. Loreal posterior to nasal, narrower, and diagonally oriented; contacting posteriorly the first superciliary and first supraocular, a preocular, and a frenocular. Frenocular small, below preocular, followed posteriorly by four
(MZUSP 96093). Scale for each 5 1 mm.
suboculars, the first three elongate, the fourth wider. Seven supralabials, fourth under the center of the eye, fifth the largest contacting fourth subocular, second the smallest; seventh separated from the granules surrounding anterior margin of the ear by an enlarged scale. Three superciliaries, first largest, wider anteriorly, longer than first supraocular, contacting preocular, loreal, first and second supraoculars, second superciliary and upper eyelid; second superciliary the smallest. An enlarged quadrangular scale follows the third superciliary. Central part of eyelid with a semitransparent undivided disc surrounded by small and slightly pigmented granular smooth scales. Lower eyelid with seven strongly pigmented palpebrals. A small postocular between quadrangular scale following third superciliary and fourth subocular. Temporal region with smooth and juxtaposed scales, irregular in size and shape, between parietals and supralabials; the larger with about the same size the fifth supralabial. Ear opening surrounded by a series of very small and juxtaposed rounded granules; external auditory meatus large, tympanum distinct, subovoid. Lateral surface of neck with 10 transverse rows of smooth, slightly imbricate and rhomboid enlarged granules. All head scales smooth and juxtaposed with scattered sensorial organs.
Mental broad, wider than high. Postmental heptagonal, wider than long. Three pairs of genials, all contacting infralabials; the first smaller, third largest, first and second in broad contact at midline; third pair contacting only anteriorly. Two series of enlarged but much smaller and symmetric, flat, and chevronlike pregular scutes follow third pair of chin shields preventing their contact posteriorly. Six infralabials, third the largest. Gulars smooth, imbricate, rounded posteriorly, in seven transverse rows; third to sixth rows with some enlarged scales that are wider than long, other scales much smaller, longer than wide. Gulars decreasing progressively in size towards side of neck. A distinct interbrachial region with nine scales that are longer than wide, elongate, smooth, and longer than the gular scales. Collar fold absent.
Dorsal scales imbricate and disposed in regular transverse rows; smooth, isodiametric or subrectangular, and rounded posteriorly in occipital region, becoming progressively narrower, lanceolate, strongly keeled, with sides almost juxtaposed. Thirty-six transverse rows of dorsals between interparietal and the posterior level of hind limbs. Lateral scales as long as, slightly wider than, and more diagonally disposed than dorsals, keeled, strongly imbricate laterally. A distinctive area with small, smooth, and rounded granules surrounds the area of arm insertion. Thirtyeight scales around midbody. Ventral scales smooth, longitudinally imbricate, laterally juxtaposed, longer than wide, rounded posteriorly, in six longitudinal and 21 transverse rows from interbrachials (excluded) to preanals. Six scales in precloacal region, central and paramedials the largest. Total pores 10, opening in the center of scale, continuous, with no gap between femoral and preanal ones.
Scales of tail smaller than midbody dorsals, otherwise identical to them; keeled, lanceolate, strongly imbricate longitudinally; those near the base of the tail larger, but in the posterior and ventral part of tail, becoming gradually identical around tail.
Forelimbs with large, smooth, and imbricate scales; those from ventral part of brachium and forearm much smaller, rounded, rhomboid, juxtaposed. Anterior and ventral parts of hind limbs with irregularly large, smooth and imbricate scales, identical to the corresponding parts of the forelimbs. Posterior part of hind limbs with granular, juxtaposed scales, grading progressively to larger, imbricate and keeled scales on dorsal part of tibia. Carpal and tarsal scales large, imbricate; supradigital lamellae smooth, imbricate. Palmar and plantar surfaces with smooth, small granules; infradigital lamellae single, 11 on finger IV and 17 on toe IV. Toes and fingers, except for finger I, clawed, and respectively in the following relative sizes: 1, 2 5 5, 3 5 4, and 1, 2, 5, 3, 4.
Dorsal surfaces of body and tail and lateral part of tail dark brown with an irregularly distributed darker punctuation generally between some dorsal scales. A cream to white longitudinal line, more conspicuous anteriorly, extends paradorsally from the occipitals to the anterior third of the tail. Flanks darker, strongly mottled by an irregular pattern of cream to yellow spots that extend to the lateral
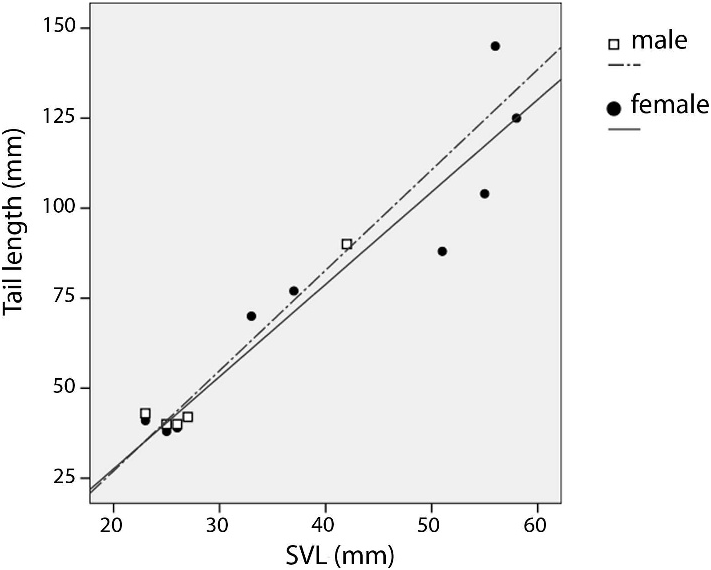
(r 2 0.91, n 5 12) of Caparaonia itaiquara from Parque Nacional do Caparaó , Brazil .
part of the tail where they become diffuse. Lateral parts of head with a similar pattern; irregular black blotches concentrated in the central parts of the supra and infralabials, yellow on their sutures. Ventral parts of body and tail predominantly cream to yellow with irregularly black spots concentrated in the central part of the scales. Ventral part of tail darker near the extremity. Limbs dark brown dorsally, irregularly mottled with a cream to yellow pattern similar to that of flanks; ventrally, cream to yellow with dark pigmentation in the center of the scales.
Hemipenis almost totally everted at preservation except for the apex; bilobation not evident; sulcus spermaticus apparently single, edged by an enlarged naked area. A series of 10–12 series transverse chevronlike, undulate, and continuous naked flounces on the opposite side of the organ. The 5–6 apical flounces end in almost free and distally bifurcate protuberances. Spines absent inside or outside flounces.
MEASUREMENTS OF THE HOLOTYPE: Snoutvent length: 53 mm; tail length: 100 mm (tip broken).
VARIATION: Males are slightly smaller and have slightly longer tails than females: maximum SVL for males and females was respectively 55 mm and 60 mm. Tail length varied respectively in males and females from 1.53 to 2.14 and 1.50 to 2.58 times SVL (r 2 males 0.94, n 5 5; r 2 females 0.91, n 5 12; fig. 3). No sexual differences were found in squamation. Variation in meristic characters (n 5 24) was the following (mean and standard deviation, respectively, in the parenthesis): dorsal rows, 36–40 (37.9 ± 1.19); ventrals 21–27 (23.9 ± 1.41); scales around midbody 36–39 (37.5 ± 0.94); infradigital lamellae under finger IV 10– 14 (11.5 ± 1.03); and infradigital lamellae under toe IV 15–20 (16.9 ± 1.37). All specimens show 3 supraoculars as well as 4 superciliaries except for MZUSP 95061 View Materials , 97619 View Materials , and 96088 where only 3 superciliaries are present. All the specimens have 7 supralabials and 7 infralabials except for MZUSP 98055 View Materials , which has 8 supralabials, and MZUSP 95061 View Materials , which has 6 infralabials. The distinctively enlarged pair of occipitals may be separated at midline by 1 or 2 scales. Gular rows vary between 7 and 8. Total number of pores varies between 9 and 10 in males and 2– 5 in females. Femoral pores are present only in males, preanal pores are present both in males and females but are much more conspicuous in males. Pores are highly conspicuous, placed in distinctively elevated scales, aligned on each side without gaps between preanal and femoral .
1 mm.
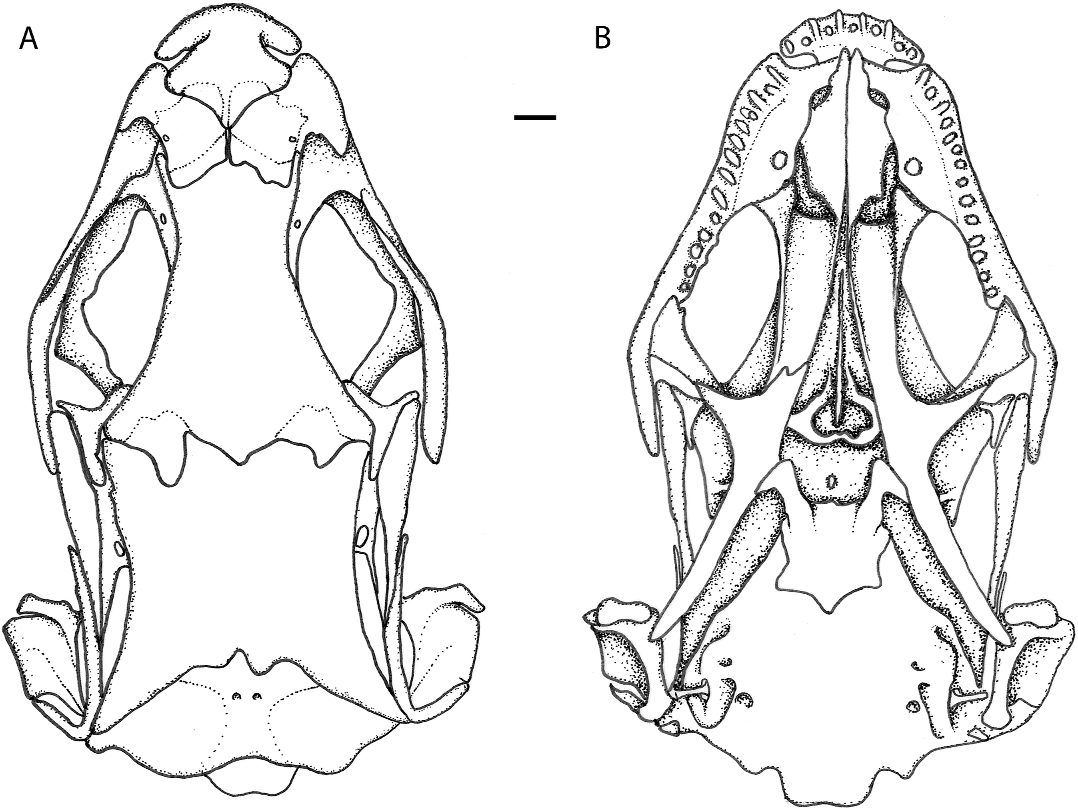
OSTEOLOGY (figs. 4 and 5): The following description is based on three alizarin-prepared skeletons (MZUSP 95054, 97619, and 97620). Premaxillary as long as wide, touching but not articulating laterally with the maxillary. Dorsal lamina of premaxillary triangular posteriorly, long, covering slightly anteriorly the nasals and indenting deeply their suture preventing their anterior contact. Eleven premaxillary conical teeth. Nasals large, slightly longer than wide, wider anteriorly, diagonally disposed, widely separated anteriorly, in midline contact in posterior third, covering anteriorly the frontal. Frontal longer than wide, strongly constricted between orbits, wider posteriorly, covering anteriorly the parietal and articulating laterally with it by a pair of frontoparietal tabs. Parietal longer than wide, wider and concave posteriorly, covering laterally the occipital region. Lateral expansion of parietal absent leaving the supratemporal fenestra open. Epipterygoid contacting externally a descending epipterygoid process of the parietal. Maxillary contacting dorsally nasal, lateral parts of frontal and lacrimal without overlapping, and covering extensively prefrontal and jugal; 14 maxillary teeth. Prefrontal large, its posterior process long but not reaching the level of middle of orbit; in broad contact with frontal. Lacrimal small, rod shaped, very conspicuous, contacting prefrontal and maxillary along the inferoanterior part of orbit. Postfrontal and postorbital single. Postfrontal roughly triangular, contacting jugal, frontal, postorbital, and parietal, closing posteriorly the orbit. Posterior part of postfrontal wider, longer, almost straight, preventing contact between frontal/parietal and postorbital and covered by the postorbital. Postorbital long and narrow, not expanded, contacting posteriorly squamosal and leaving supraorbital fenestra widely open. Squamosal long, posteriorly curved and fitting at the top of the quadrate. Supratemporal fenestra widely opened, not constrained laterally by lateral expansions of parietal and postorbital, which are absent. Supratemporal present, small, in straight contact with posterior part of parietal and squamosal. Fifteen scleral ossicules in the eyeball. Vomer, palatine, pterygoid, and ectopterygoid present. Vomer, palatine, premaxillary, and maxillary in contact restricting the fenestra exochoanalis. Infraorbital fenestra large, bordered posteriorly by ectopterygoid and pterygoid. Pterygoid teeth absent. Stapes rodlike, wider, and rounded at base. Sutures between supraoccipital, exoccipital, basioccipital, and the otic area of the skull are not well visible in the articulated skeleton, nor are those eventually between basioccipital and basisphenoid.
In the lower jaw dentary, articular, splenial, angular, and suprangular are distinct; there are 17 dentary teeth, conical anteriorly, bicuspid or tricuspid posteriorly.
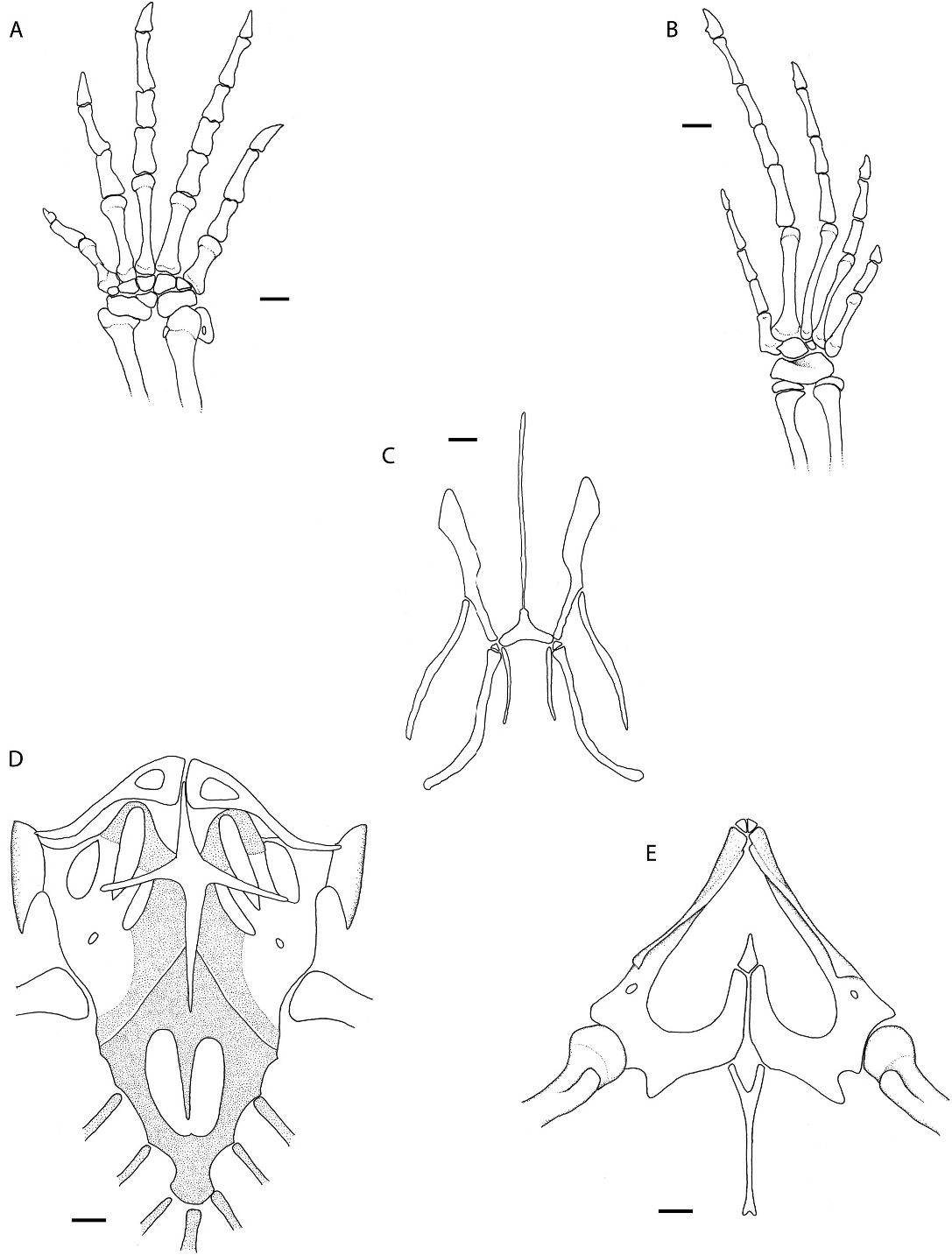
Glossohyal long, fused to basihyal. First ceratobranchial curved posteriorly; hypohyal and ceratohyal present. A second short pair of ceratobranchials is present and is positioned parallel to the anterior part of the trachaea.
Anterior part of clavicle very enlarged, flattened, enclosing a fenestra. Interclavicle long, cruciform, with very long lateral processes reaching the sternum but not sternal fenestra. Scapulocoracoid with coracoid, scapular, and scapulocoracoid fenestrae; suprascapula present. Sternum with a large fenestrum, and receiving three sternal ribs and a xiphisternum with two inscriptional ribs. Ilium, ischium, and pubis present, the latter with a conspicuous pectinate apophysis. Hypoischium long, wider at the base, almost reaching the preanal border; a small elongate preischium and a small quadrangular ossified prepubis are present.
Thirty procelous presacral vertebrae; neural spines low, higher anteriorly; hipapophyses present on first eight vertebrae; zigantrumzygosphene present. Last presacral vertebra lacking ribs. Two sacral vertebrae. First four caudal vertebrae lacking autotomic processes, with long and wide transverse processes, and with wide and high neural spines. From the fifth vertebra on intravertebral autotomic septa are present, transverse processes are narrow and neural spines become lower.
Humerus and femur slightly larger than radius and ulna and tibia and fibula. Remaining elements of fore- and hind limbs as in figure 5.
HABITAT, DISTRIBUTION, AND NATURAL HISTORY
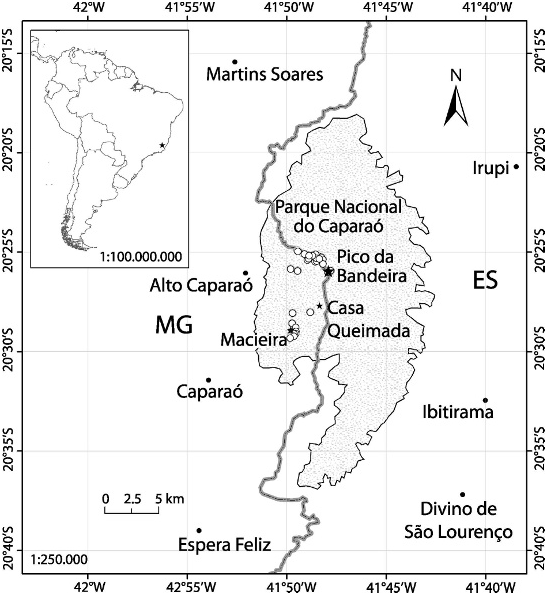
Caparaonia itaiquara is currently known only from and considered endemic to the mountains of Parque Nacional do Caparaó, an area with 168,000 km 2 near the border of the states of Minas Gerais and Espirito Santo (fig. 6), Brazil. This highly elevated and deeply dissect- ed mountainous area is formed by middle to late Tertiary uplifted blocks of igneous or highgrade metamorphic rocks and includes the three highest mountains of eastern tropical South America: Pico da Bandeira (2890 m), Pico do Calçado (2840 m), and Pico do Cristal (2780 m), respectively the third, fourth, and sixth highest mountains in Brazil ( Safford, 1999a, 1999b). The high elevation, plus their exposure to southern polar fronts, makes these mountains the coldest spots in eastern South America. These conditions confer to this area a cool and humid climate, and patterns of rainfall, temperature, and frost that are distinctly seasonal ( Safford, 1999a). Dry season extends from March to September and rainy season extends from November to January; annual mean precipitation varies between 1250 and 1500 mm; June and July are the coolest months, December the hottest. There are no climatic stations in the highest mountains, but according to IBAMA (Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis), the Brazilian environmental agency, the maximum and minimum annual temperatures vary respectively from 34 ° to 36 ° C and 24 ° to 0 ° C. Estimated annual mean temperatures calculated after regional elevational temperature gradients (ca. 0.55 ° C/ 100 m) and verified by soil temperatures measured at less than 70 cm indicates that annual mean temperatures fluctuate around 10.5 ° C at 2400 m, and 8.5 ° C at 2800 m ( Safford, 1999b). We left a data logger at 2400 m altitude, at a relatively protected site near Pico da Bandeira, between December 2004 and November 2005. It registered an annual average temperature of 11.1 ° C, a minimum temperature of 2.5 ° C in July, and a maximum of 30.7 ° C in December. The daily mean temperature range varied around 2 ° –3 ° C throughout the year, with the hottest hours of day happening from midday to 4:00 PM (fig. 7). The wider range for December 2004 reached 6 ° C, but it was probably due to the fact that temperatures were recorded for four days only.
Two main habitat types occur at Parque Nacional do Caparaó. The Atlantic rainforest and a less diverse and gradually lower montane forest are the dominant physiognomy up to about 1800–2000 m elevation. These forested habitats were severely affected by fires and the expansion of coffee plantations in the past and are presently restricted to a few isolated spots. Above 2000 m the Caparaó massif is characterized by open and rocky areas dominated by shrubs and grasses referred to as campos de altitude or campos rupestres (fig. 8A–D). They are currently isolated open areas on top of the Atlantic forest domain. Landscape in these open areas consists of roughly rounded hilltops, plateaus of variable extension and deeply dissected valleys with stream courses. The entire area is characterized by the presence of rocky outcrops or considerable extensions of bare rock; high cliffs are also very typical in areas of sharp relief. Soils are humid and dark colored (Modenesi, 1988) and support a diverse flora characterized by a high degree of endemism. Local vegetation, like tussock grasses, rosetteforming plants, sclerophyllous shrubs, and ground-hugging cushion plants, are morphologically similar to those found in most tropical highland environments with low mean temperatures, diurnal freeze-thaw cycles, and relatively moderate seasonal variations in temperature ( Smith and Young, 1987; Smith, 1994). Shrubs (especially species of Baccharis View in CoL , Vernonia View in CoL , various Eupatorieae, Tibouchina View in CoL , Leandra , and Myrtaceae View in CoL ) and small copses of short, often stunted trees (e.g., Escallonia View in CoL , Weinmannia View in CoL , Rapanea , Symplocos, Maytinus View in CoL , Roupala View in CoL ) are common and coexist with a more continuous coverage of buchgrasses ( Cortaderia View in CoL , Calamagrostis View in CoL , Andropogon View in CoL ) and large thickets of bamboo ( Chusquea sp. ). At some places a sparse understory of herbs and pteridophytes might be present. There is almost no zonation of the vegetation in response to elevation ( Safford, 1999a).
All specimens of Caparaonia itaiquara were found in the highland open areas between 1873 m and 2770 m elevation from 9 AM to 6:45 PM. Figure 6 View Fig shows a map of the area with all localities where specimens were seen. Of the 25 specimens observed and apparently resting, 22 were found under rocks, two under tree trunks, and one inside a small termite mound. Only one specimen was moving actively among tickets of Chusquea and other vegetation at 2:15 PM. Twenty (90%) of the 22 specimens obtained under rocks were under rocks situated on extensive rocky outcrops, the other two under isolated rocks on loose soil with grasses and small bushes. The termite mound that was sheltering one specimen was built between a small rock and an extensive rocky floor. We also found eggs and egg shells, all under rocks situated in extensive rocky floors. Rocks sheltering specimens and/or eggs varied in dimensions from 0.25–1.43 m long X 0.22–0.59 m wide X 0.14–0.33 m high; mean dimensions for length, width, and height were respectively, 0.65 m, 0.42 m, and 0.22 m. Underneath these rocks a thin, soft, and generally humid soil layer, almost always with roots of the surrounding vegetation, was usually present. In some cases part of the rock was in direct contact with the rock outcrop. Lizards were generally seen immobile, immediately after the rocks were overturned, inside small and irregular soil cavities connected to superficial galleries. Eggs/eggshells were always found in pairs (minimum of 2) in small cavities under rocks. When more than two eggs or eggshells were discovered, the shell condition clearly indicated that they could be assembled in pairs and these pairs attributed to different clutches probably laid at different times. The maximum number of developing eggs found under a rock was four, the two pairs clearly laid at different times; the maximum number of eggs/eggshells observed under the same rock was 10. In one case, the clutch consisted of 2 developing eggs plus 4 pairs of eggshells; in the other, 5 pairs of eggshells were found beside an adult female. These observations suggest that, as usual in microteiids, clutch size in Caparaonia itaiquara is limited to two eggs, and that egg-laying sites are frequently reused. Our data are also suggestive that communal nests occur under the same rock and/or females return to the same place to lay their eggs. As we have almost no life-history data on these lizards, we cannot give a conclusive answer to these questions. The first hypothesis seems more probable considering the climate of unsuitable conditions for multiple clutches per season due to the restricted period of reproductive activity of these high-altitude lizards and their probably short life expectancy.
Ants, beetles, spiders, earthworms, or myriapods were sometimes found near the lizards or inside the small soil cavities where they frequently take refuge, and may serve as their prey. The only other lizard occurring with Caparaonia itaiquara in the open areas in the highlands of Parque Nacional do Caparaó is an isolated population of the viviparous skink that we provisionally identify as Mabuya dorsivittata , which is active on sunny days on the ground and on thickets of vegetation.
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Caparaonia itaiquara
| Rodrigues, Miguel Trefaut, Cassimiro, José, Pavan, Dante, Curcio, Felipe Franco, Verdade, Vanessa Kruth & Pellegrino, Kátia Cristina Machado 2009 |
Caparaonia itaiquara
| Rodrigues & Cassimiro & Pavan & Curcio & Verdade & Pellegrino 2009 |
Leandra
| Raddi 1820 |