
Calotes zolaiking, Giri & Chaitanya & Mahony & Lalrounga & Lalrinchhana & Das & Sarkar & Karanth & Deepak, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4638.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:182957CC-29AC-4E8D-B66F-8C04B5561F92 |
|
DOI |
https://doi.org/10.5281/zenodo.4324181 |
|
persistent identifier |
https://treatment.plazi.org/id/03B087FB-C43D-FF90-FF17-BBDCFC04FDB0 |
|
treatment provided by |
Felipe |
|
scientific name |
Calotes zolaiking |
| status |
sp. nov. |
Calotes zolaiking View in CoL sp. nov.
Figs 8–12 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 ; Tables 3–5.
Holotype. Adult female, NCBS-AU152 , from Synod Hospital Compound (23°46 19.06 N, 92°43 56.37 E, 1290 m a.s.l.), Durtlang, Aizawl District , Mizoram state, Northeast India, collected by C. Lalrinchhana and Samuel Lalrounga on 10 June 2014.
Paratypes. Two subadult females, NCBS-AU153 and ESV 105, an adult female, NCBS-AU154 , and a subadult male, NCBS-AU155 , collection data same as holotype; subadult male, BNHS 2327 View Materials , from Hmuifang (23°27 17.93 N, 92°45 07.05 E, 1478 m a.s.l.), Aizawl District , Mizoram state, Northeast India, collected by C. Lalrinchhana and Samuel Lalrounga.
Referred specimens. Adult male, NCBS-AU156 collection data same as holotype .
Diagnosis. A medium sized Calotes , snout to vent length averaging 64.7 ± 8.17, and maximum to at least 77.0 mm. Body feebly compressed laterally with a weak dorsal crest; scales on top of head highly heterogeneous, keeled, those above orbits are largest, scales surrounding parietal are unequal in size; three spines on each side of head, one above orbit, one above tympanum and one on temporal region; dorsal scales heterogeneous, composed of medium sized, weakly pointed, strongly keeled scales, intermixed with similar but slightly larger scales which are distinct on flanks, upper rows directed backwards and upwards and a few lower rows directed backwards; 49–52 midbody scale rows; a weakly developed fold anterior to forelimb insertion having granular scales; tympanum small; tail rounded; eight to nine supralabials and seven to ten infralabials; lamellae bicarinate, 21–24 on fourth finger and 23–27 on fourth toe.
Calotes zolaiking sp. nov can be easily diagnosed from all congeners except Calotes paulus comb. nov. in possessing heterogeneous scales on the dorsum and a weakly developed dorsal crest. Based on dorsal pholidosis, the new species is most similar to Calotes paulus comb. nov., but differs with respect to the following ( Calotes paulus comb. nov. given in parentheses): greater number of midbody scales rows, 49–52 (versus 42–46), greater number of caudal vertebrae, 50 (versus 35–45), and fewer caudal vertebrae with transverse processes 12 (versus 14).
Description of holotype (NCBS-AU152). Specimen is generally in good condition but slightly dehydrated, tail bent toward left in a sigmoid manner, left manus slightly adpressed with outer three fingers curved, right manus ad- pressed with fingers directed backwards, eyes sunken, orbital spine on left side broken, tips of supra-tympanic spines on both sides curved, all artefacts of preservation. Mensural and meristic data is summarised in Table 3. An adult female, SVL 77.2 mm. Head relatively long (HL/SVL 0.31), broad (HW/HL ratio 0.65), not depressed (HH/HL ratio 0.52), noticeably broader than neck ( Fig. 8A View FIGURE 8 ). Snout short (SE/HL ratio 0.37), longer than orbit diameter (OD/SE ratio 0.78). Orbit large (OD/HL ratio 0.29); pupil round, eyelids covered with small rounded scales, a single row of scales bordering eyelids slightly elongate, six supraciliaries on each side, elongate, subimbricate, slightly protruding laterally on supraorbital ridge, similar in size, with single anterior most supraciliary smallest. Snout obtusely pointed; rostral wider (2.1 mm) than deep (0.7 mm), contacted laterally by first supralabial, an elongated large prenasal, and two large scales dorsally. Canthus rostralis and supraciliary edge sharp. Nostril circular, laterally positioned and placed at centre of a large, undivided nasal plate ( Fig. 8C View FIGURE 8 ), which is bordered by seven scales on both sides, including one prenasal, one supranasal, three postnasals, with two subnasals on left side, and one subnasal and second supralabial on right side; separated on both sides from rostral and first supralabial by prenasal. Supralabials roughly rectangular, more or less equal sized, posterior-most being longest, bordered above by two rows of scales starting behind postnasals and extending to posterior border of orbit; upper row distinctly enlarged consisting of equal sized, roughly rectangular scales; scales on lower row are heterogeneous in shape and size, decreasing in size posteriorly. Loreal region concave, scales of loreal region heterogeneous in shape and size, flat or weakly tuberculate. Scales on postorbital and temporal region heterogeneous in shape and size, subimbricate, mostly tuberculate; a row of five much larger longitudinally keeled scales, fourth being largest, extending from posterior midorbital region to just before anterior edge of tympanum ( Fig. 8C View FIGURE 8 ). Tympanum covered with single scale ( Fig. 8C View FIGURE 8 ). Canthals enlarged, overlapping, becoming slightly larger along supraciliaries. Scales on dorsal surface of snout and forehead heterogeneous in shape and size, all weakly pointed anteriorly ( Fig. 8B View FIGURE 8 ), smaller scales weakly tubercular, larger with distinct keels except those on internarial region which are roughly rounded, uniformly large and smooth. Supraorbital scales weakly keeled, increase in size from supraciliaries to inner edges of orbits, of which enlarged scales follow curvature of orbits posterolaterally ( Fig. 8B View FIGURE 8 ). Two small scale rows divide enlarged scales of inner orbits at narrowest point of frontal. Scales on occipital region heterogeneous in shape and size; smaller scales weakly tubercular, larger ones with distinct keels. Parietal scale roughly triangular, without visible pineal eye (but with distinct depression) ( Fig. 8B View FIGURE 8 ). Parietal scale bordered laterally by three enlarged, roughly elongated keeled scales, with much smaller scales anteriorly and posteriorly. Single temporal spine on each side, longer than nuchal spines; shorter orbital spine and a small supra-tympanic spine also present on each side ( Fig. 8B View FIGURE 8 ). Mental subtriangular, approximately as wide as long, bordered posterolaterally by a row of six elongated postmental chin shields on either side, gradually changing shape and size posteriorly to blend with surrounding rows of gular scales ( Fig. 8D View FIGURE 8 ). First pair of postmental chin shields in contact laterally with first infralabials, remaining shields separated from infralabials by one to two rows of small gular scales. Posterior gular region with considerably enlarged, flat, strongly keeled, pointed, imbricate scales, those on jowls slightly larger in size; anterior gular scales much smaller, rounded, weakly keeled and subimbricate; all gular scales directed posteromedially except a few median rows which are directed posteriorly. No distinct gular pouch present, transverse gular fold absent ( Fig. 8D View FIGURE 8 ). Ventral scales are like those on posterior gular region, enlarged, flat, strongly keeled, pointed, imbricate, homogenous in shape but heterogeneous in size, generally increasing in size laterally; arranged in regular longitudinal rows that are directed posteriorly, but those on lateral aspect are directed posteromedially ( Fig. 9C View FIGURE 9 ). Approximately 49 scales around mid-body. Nuchal crest composed of three short, laterally compressed, equal sized spines. Remaining vertebral scales slightly enlarged relative to adjacent rows ( Fig. 9A View FIGURE 9 ) but possess a more pronounced median keel providing a serrated appearance in profile ( Fig. 9B View FIGURE 9 ); 33 mid-dorsal scales from first raised nuchal scale to above level of cloaca. Dorsal scales heterogeneous in size and shape; all scales with a moderate median keel, arranged into approximate rows ( Fig. 9A & B View FIGURE 9 ); keels on those of upper flanks mostly oriented obliquely upward, horizontal on mid-flanks and obliquely downward on lower flanks ( Fig. 9B View FIGURE 9 ). A distinct shallow oblique fold in front, and curving around anterior forelimb insertion. Scales of forelimbs and ventral hindlimbs form approximate rows, those on dorsal hindlimbs do not form regular rows and are heterogeneous in size. Dorsal scales larger than ventral scales on forelimbs and hindlimbs, all moderately keeled dorsally, very weakly keeled ventrally. Forelimbs moderately long (UAL+LAL+HaL/SVL ratio 0.45); hindlimbs long (ULL+LLL+FoL/SVL ratio 0.66). Digits slender, elongate and ending in a strong, slightly curved claw. Lamellae entire, bicarinate; 21 on fourth finger, 23 on fourth toe. Relative finger lengths: IV(8.5 mm)> III(7.9 mm)> II(4.6 mm)> V(4.1 mm)> I(3.0 mm) and toes IV(11.9 mm)> III(10.5 mm)> V(8.2 mm)> II(7.2 mm)> I(5.5 mm). Tail entire, rounded, nearly twice head-body length (TL/SVL ratio 1.87), not swollen at base ( Fig. 8A View FIGURE 8 ); uniformly covered with similar sized, strongly keeled, weakly pointed, regularly arranged, posteriorly directed imbricate scales.
Coloration (in preservative). Coloration in preservation is more or less similar to live specimens but slightly faded and predominantly light brown. Transverse stripes on body, limbs and tail are darker. Refer to Figures 8 View FIGURE 8 and 9 View FIGURE 9 for further details of coloration and markings.
Coloration in life (based on specimens other than the holotype). Coloration in life varies, but is primarily mottled shades of light and darker brown dorsally, with four distinct lighter transverse stripes on body, two on forelimbs and two on hindlimbs; stripes may be primarily green or light brown ( Fig. 10 View FIGURE 10 ). Two thin white longitudinal stripes on upper flanks extend from rear of head to base of tail. Region below orbit and lateral surface of snout white, with narrow white and brown radial stripes extending from eye. Tail with faint light and darker brown bands. Ventral surfaces whitish, throat with irregularly arranged dark mottling or thin stripes. Refer to Figure 10 View FIGURE 10 for further details of coloration and marking on live animals.
Variation. Paratypes and referred specimen agree with holotype in general morphology and scalation with following exceptions: NCBS-AQ154 has ten infralabials on both sides, and ESV 105 has seven infralabials on both sides; 35 vertebral scales on BNHS 2327; within paratype series, lamellae on fourth finger varies from 22 to 24, and on fourth toe from 25 to 27. Further variation in mensural and meristic characters within paratype series is documented in Table 3.
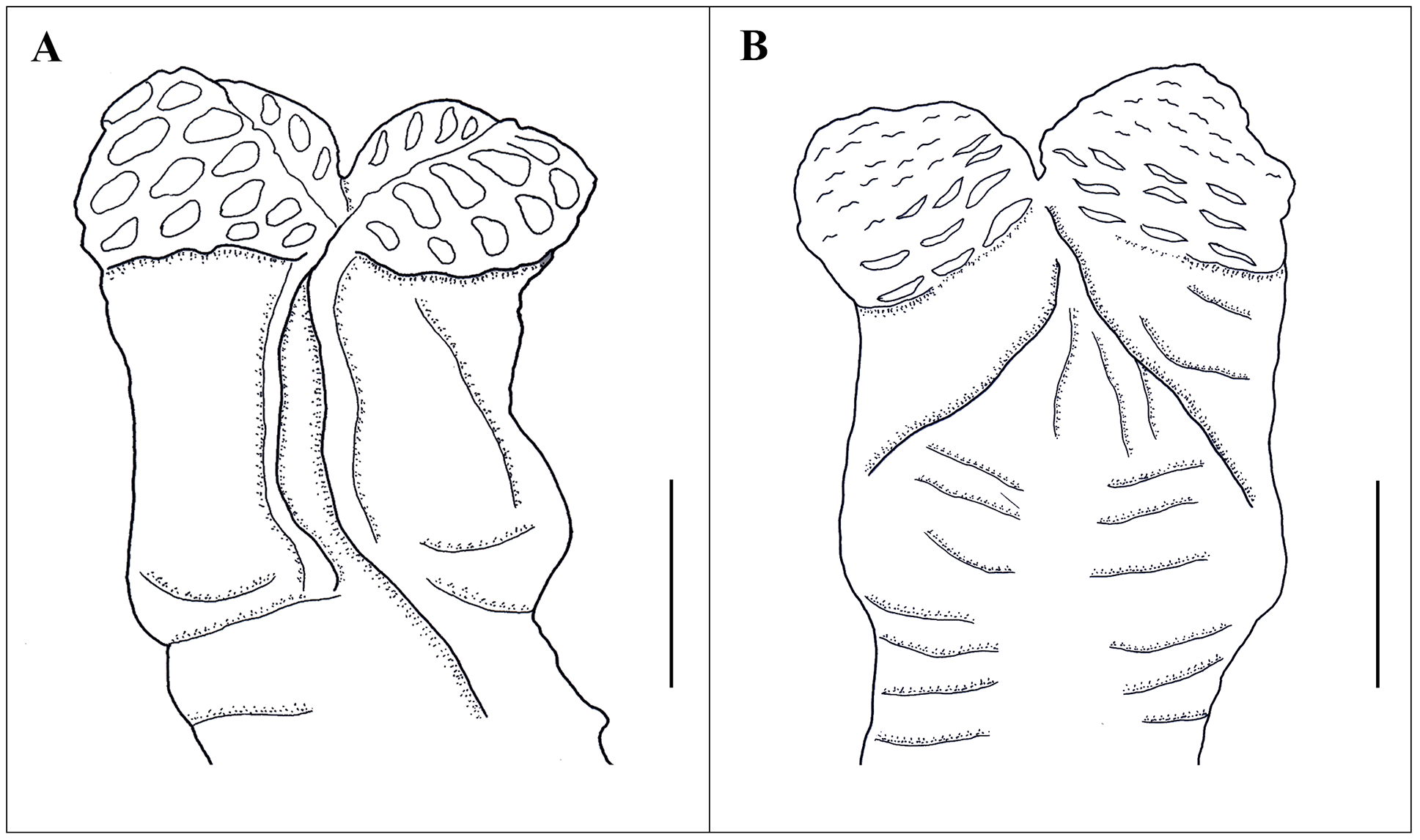
Hemipenis. Hemipenis single, clavate (divided less than half of length) with length of organ greater than width. Base naked; sulcus spermaticus broad, canal like, shallow at base and deeper at terminal region. Lips of sulcus spermaticus smooth. Calyculate ornamentation present on each lobe. Thick walled smooth calyces form deep oval pits. Fleshy cardioid structure at base of ventral sulcus absent. Apex capitate and divided into four segments ( Fig. 11 View FIGURE 11 ). Hemipenis of Calotes zolaiking sp. nov. matches with 13 out of 14 characters reported for seven other Calotes spp., and 12 out of 14 characters reported for Calotes ceylonensis Müller, 1887 ( Maduwage et al. 2008; Table 4).
Osteology. Skull of Calotes zolaiking sp. nov. (NCBS-AU156) is subpentagonal in outline ( Fig. 12A View FIGURE 12 ): raised in parietal region and slopes steeply towards naris ( Fig. 12B View FIGURE 12 ). Hyoid apparatus broad with a paired ceratobranchial (CB), CB II shorter than CB I. Maxilla has 14 teeth (including one canine-like tooth [on each side]); three premaxillary teeth; thirteen teeth on either side of mandibles ( Fig. 12D & E View FIGURE 12 ). Twenty presacral vertebrae excluding atlas and axis and 50 caudal vertebrae ( Table 5). Sternum with an oval and elongated central foramen. Ten trunk vertebrae have ribs, and an additional five vertebrae near pelvic girdle have ribs replaced by small transverse processes. Shoulder girdle comprises a broad clavicle; interclavicle long and rod like, suprascapula wedge shaped. Humerus with well-developed proximal and distal ends. Phalangeal formula of manus is 2:3:4:5:3 and of pes is 2:3:4:5:4. Many osteological characters checked for Calotes zolaiking sp. nov. (NCBS-AU156) matched those of Calotes paulus comb. nov., except the former had a higher number of caudal vertebrae ( Table 5). Calotes paulus comb. nov. and Calotes zolaiking sp. nov. differ from other congeners in five different osteological characters ( Table 5). Calotes paulus comb. nov. and Calotes zolaiking sp. nov. differ from Pseudocalotes in three different osteological characters ( Table 5). Within the genera Calotes and Pseudocalotes , there are several variable characters ( Table 5).
Etymology. The species is named for its occurrence in high elevation regions. The specific epithet is a noun in apposition, derived from the Mizo language and is a portmanteau word for a lizard that inhabits high elevations (Mizo: Zo = highland/cold region, Laiking = agamid lizard).
Suggested English name. Mizoram Montane Forest Lizard .
Distribution and natural history. Calotes zolaiking sp. nov. is known only from its type and paratype localities, Durtlang (1290 m a.s.l.) and Hmuifang forest (1480 m a.s.l.) in Aizawl District, Mizoram state, Northeast India. These two localities are approximately 36 km apart [straight line distance]. This species is also suspected to be found in other high elevation areas of Mizoram. However, very few herpetological surveys have been carried out in this state, so the full extent of its distribution and preferred elevational range is currently unknown. The holotype was collected from a residential area where both native and non-native trees were present. It was seen perching on a Melaleuca citrina (Curtis) Dum.Cours. (bottlebrush tree), an introduced ornamental species native to Australia. The native trees dominating the locality include Trema orientalis Blume , Callicarpa arborea Roxb. , Schima wallichii (DC.) Korth., Castanopsis tribuloides Sm. , Albizia chinensis (Osbeck) Merr. and Ficus semicordata Buch. -Ham. ex Sm. The locality where the paratypes were collected is dominated by Rapanea capitellata (Wall) Mez. , a Eurya sp. Korth., Quercus spp. L., Elaeocarpus rugosus Roxb. , Nyssa javanica (Blume) Koord. , a Macropanax sp. Miq., Schima wallichii (DC.) Korth. and Ardisia macrocarpa Wall. , with a small patch of grassland ( Fig. 13 View FIGURE 13 ). According to Champion and Seth (1968), these regions fall under their “Montane Sub-tropical Forest” category. Most of the natural history observations of Calotes zolaiking sp. nov. were made from Hmuifang where the paratypes were collected. The type series was collected from vegetation at a height of 1.5 – 4.5 m above ground level. All individuals of the species were spotted perching on branches, which suggests they are primarily arboreal. In captivity, an adult female (NCBS AU154) laid three large eggs (measuring 14.1 X 8.2 mm, 14.3 X 8.5 mm and 14.5 X 8.5 mm) in July. During fieldwork at the type locality, juveniles were observed in November. Other agamids found in sympatry include Calotes cf. versicolor and Cristidorsa otai ( Mahony, 2009) .
Phylogenetic relationships and morphology. Based on its phylogenetic position within the subfamily Draconinae , Oriocalotes now becomes the second monotypic genus (after Brachysaura : see Deepak et al. 2015) in recent years to be subsumed into Calotes . The poor support in the nuclear dataset is probably due to incongruence in the individual nuclear genes (Appendix 4–6). So, additional nuclear markers need to be analysed to further resolve its position within the Calotes radiation. Our nuclear and mitochondrial trees support a clade encompassing Calotes + “ Oriocalotes ” + Psammophilus . Members of this radiation are largely confined to India and harbor multiple ecomorphs; Psammophilus is a predominantly rock dwelling genus with a dorsoventrally compressed body. Calotes minor is completely terrestrial with a relatively short fifth toe ( Deepak et al. 2015), whereas all other Calotes (including Calotes paulus comb. nov. and Calotes zolaiking sp. nov.) are predominantly or exclusively arboreal and have a longer fifth toe.
Mahony (2010) noted that Oriocalotes shares common external morphological characters with Calotes (elongated spines on the post ocular, temporal and supratympanic regions) and Pseudocalotes (enlarged subocular scale row and heterogeneous dorsal scalation). Mahony (2010) also noted that no Calotes species possessed a noticeably enlarged subocular scale row, however, this character was found to be present or absent in the genus Diploderma Hallowell, 1861 , so the plasticity of this character within Calotes (now including Oriocalotes ) might not be unusual. The evolutionary relevance of this character is yet to be studied. Hallerman and Böhme (2000) (modified by Deepak et al. 2015) attribute the following characters to the genus Calotes : a broad head (HW/HL ratio 0.58–0.82), groups of spines from eye to tympanum, cheek often swollen in males, relatively long hindlimbs (hindlimb length/SVL ratio 0.62–1.01) and tail (TL/SVL ratio 0.9–3.0). Presence of regular, uniform dorsal scales, thought to be an important character in the diagnosis of the genus Calotes ( Smith 1935; Manthey & Denzer 2000), is now demonstrated to have no systematic importance, as observed in the draconine lizards belonging to the genus Bronchocela Kaup, 1827 and Pseudocalotes .
Our study reinforces the importance of hemipenial morphology as a generic diagnostic character for agamid lizards. The shape of the hemipenis is largely conserved within draconine genera ( Maduwage et al. 2008; Maduwage & Silva 2012), however, the absence of the cardioid structure on the hemipenes of Calotes zolaiking sp. nov. warrants a relook at the usage of this character to diagnose the genus Calotes ( Maduwage et al. 2008; Maduwage & Silva 2012; Deepak et al. 2015).
The osteological characters suggested by Moody (1980) to diagnose agamid genera will require revision considering the recent changes in agamid taxonomy, e.g. within the distantly related genus, Pseudocalotes , there are multiple osteological characters which are not congruent with Moody’s (1980) diagnoses ( Table 5). However, osteological comparisons of additional species in the genus Calotes is required to better understand homologous osteological characters. The genus-level assignment of draconine species has been historically based on synapomorphic characters. Taxonomic stability for most draconine species will only occur following detailed molecular phylogenetic analysis and a thorough reassessment of morphological characters.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.