
Bythinella angelitae
|
publication ID |
https://doi.org/ 10.5281/zenodo.178388 |
|
DOI |
https://doi.org/10.5281/zenodo.5629037 |
|
persistent identifier |
https://treatment.plazi.org/id/03E2F349-FF9C-7403-FF0C-CF6FFF44FE80 |
|
treatment provided by |
Plazi |
|
scientific name |
Bythinella angelitae |
| status |
|
Bythinella angelitae nom. nov.
Synonymy: Paludinella opaca Ziegler — Frauenfeld, 1857: 576, fig. 6, non Gallenstein, 1848: 18–19.
Type material: Lectotype of Paludinella opaca Frauenfeld, 1857 , here designated, and holotype of Bythinella angelitae : NHMW (Naturhistorisches Museum Wien) 103753 (Veldes See = Lake Bled, Slovenia); paralectotypes of P. opaca Frauenfeld, 1857 and paratypes of B. angelitae : NHMW 103754 (4, Veldes See = Lake Bled, Slovenia); NHMW 103758 (three specimens, no locality data); NHMW 103759 (three specimens, no locality data).
Type locality: Through our designation of the lectotype from the Slovenian sample, the type locality is restricted to Veldes See = Lake Bled, which, since the species is not lacustrine ( Radoman, 1976), must be conceived as “the area around Lake Bled”.
Additional material: spring in Tscheppaschlucht, southern Carinthia, 46.4837°N, 14.2632°E, 680 m asl., leg. P. Mildner, 30 March 2006.
Etymology: This species is dedicated to the first author’s wife Angela “Angelita” Schmitz-Ornés.
Description: Shell ( Figs 2 View FIGURE 2 , 3 View FIGURE 3 ; Table 2 View TABLE 2 ) conical to subcylindrical with typical blunt apex, holotype 3.18 mm high and 1.71 mm wide, with 4 whorls, shells from the Tscheppaschlucht 2.61 ± 0.08 mm (range 2.48– 2.76 mm) high and 1.67 ± 0.06 mm (range 1.56–1.80 mm) wide, with 3 3/8–3 7/8 convex whorls of which protoconch comprises 1.1–1.2 whorls with delicately pitted spiral lines, teleoconch without structure apart from growth lines; aperture ovate with typically pronounced angle posteriorly; umbilicus largely covered; operculum corneous, yellow, thin, ellipsoidal, paucispiral, nucleus submarginal.
Tentacles without conspicuous ciliation; epidermis black except mantle edge; ctenidium with 21–26 gill filaments, connected to pericardium with short vessel, osphradium under or slightly behind middle of ctenidium, elongate, about 1/3 or somewhat less of gill length, kidney may penetrate slightly into the mantle roof; radula formula R: 4–5 1 4–5/1-2 1–2, L: 3 1 4, M1: 17–23, M2: 27–31.
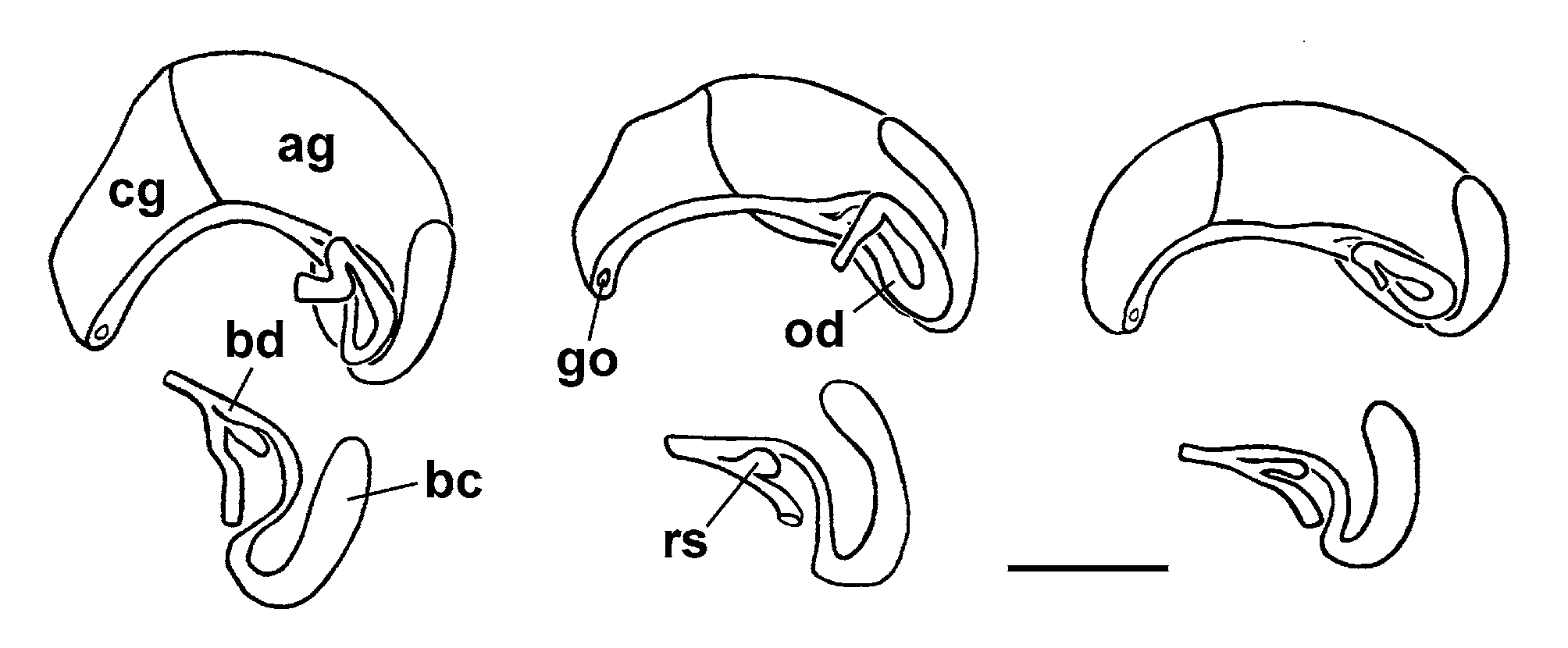
Stomach without appendix, rectum with shallow loop along pallial genital glands; ovary small sac with wide lobes comprising 0.25 whorls, starting about 1 whorl below apex, distal end close to but not reaching posterior end of stomach, albumen gland comprising three fifth length of pallial oviduct, renal oviduct describing wide loop of 270°, seminal receptacle small, tubular, arising close to junction with bursal duct, receptacle and bursal duct may be embedded into albumen gland tissue, bursa copulatrix a long sac arching around posterior end of albumen gland, bursal duct entering ventrally ( Fig. 6 View FIGURE 6 ); testis lobate, comprising about 1 whorl, starting about 0.5 whorls below apex, covering posterior chamber of stomach, seminal vesicle comprising 0.25 whorls, proximal vas deferens entering kidney-shaped prostate just in front of posterior wall of mantle cavity, distal vas deferens emerging at distal tip of prostate becoming muscular ductus ejaculatorius coiling in proximal third of resting penis, penis longer than tubular accessory gland, anterior third of penis with black pigment.
Habitat and distribution: It is only known from springs in the area of Lake Bled in Slovenia and in the Tscheppaschlucht just across the Austrian border in Carinthia.
Remarks: Morphologically and anatomically, B. angelitae nom. nov. falls well into the range of variation of its immediate sister species, B. opaca , which is also stem species of B. robiciana characterized by a globular, derived shell shape (see below). The only distinguishing feature of the new species is the denticulation of the marginal teeth of the radula, with the fewest number of denticles on the inner marginal and the highest number on the outer one. Although there is some overlap with the remaining populations — hardly though in case of the outer marginal —, the shift towards the extreme ends of the overall distribution in opposite directions must be considered distinctive. There are reports of phenotypic plasticity of radula tooth shape depending on diet ( Padilla 1998; Andrade & Solferini 2006), but we are not aware of any case of phenotypical plasticity in numbers of denticles. The numbers representing the extreme ends of the total variation observed among the populations investigated suggest that probably more than one controlling genetic locus is involved. In addition, B. angelitae nom. nov. turned out to be sister species of B. opaca and B. robiciana with HKY + I + Γ distances> 0.048 in the phylogenetic analysis based on the 638 bp long fragment of COI. The combination of these facts justifies recognition as separate species.
According to paragraph 72.7 of the ICZN the types of the species whose name is to be replaced are also types of the new name regardless of any restriction of the applicability of that name as in the present case. Therefore, the samples from Italy preserved their status as types although they most likely represent a different species (see Introduction).
Frauenfeld’s (1857) Lake Bled-sample apparently consisted of representatives of two populations judging from the color of the periostracum and the different ways of original treatment. The three lighter shells had been glued onto black cardboard (see Fig. 2 View FIGURE 2 ), whereas the two darker shells were removed from white cardboard.


| NHMW |
Naturhistorisches Museum, Wien |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |