
Brookesia brunoi, Crottini, Angelica, Miralles, Aurélien, Glaw, Frank, Harris, James, Lima, Alexandra & Vences, Miguel, 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.208594 |
|
publication LSID |
lsid:zoobank.org:pub:FF22F75B-4A07-40D9-9609-1B8D269A921C |
|
DOI |
https://doi.org/10.5281/zenodo.5617287 |
|
persistent identifier |
https://treatment.plazi.org/id/F2389052-FFB7-FF80-59E7-4137DC05F832 |
|
treatment provided by |
Plazi |
|
scientific name |
Brookesia brunoi |
| status |
sp. nov. |
Brookesia brunoi sp. nov.
( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Holotype. ZSM 888/2010 ( ZCMV 12784), adult male, collected in Anja Reserve (see Fig. 1 View FIGURE 1 in Crottini et al. 2011), Ambalavao 21°51'06.8'' S, 46°50'38.5'' E, about 950 m a.s.l., Haute Matsiatra Region, Fianarantsoa province, southern central Madagascar, on 8th December 2010 by Aurélien Miralles and Fanomezana M. Ratsoavina. A 3 mm long piece of muscle of the right hind limb has been cut and preserved in 96% EtOH for genetic analyses.
Paratypes. ZSM 889/2010 ( ZCMV 13022), adult gravid female, collected in Anja Reserve, Ambalavao 21°51'2.64'' S, 46°50'33.80'' E, 949 m a.s.l., Haute Matsiatra Region, Fianarantsoa province, southern central Madagascar, on 9th December 2009 by Angelica Crottini, D. James Harris, Iker A. Irisarri, Alexandra Lima, Solohery Rasamison and Emile Rajeriarison. One mm of the tail tip has been cut and preserved in 96% EtOH for genetic analyses. UADBA uncatalogued ( ZCMV 12783), adult female, collected at the same locality and dates of the holotype and hosted in the collection of the Université d'Antananarivo, Département de Biologie Animale. A piece of muscle has been cut and preserved in 96% EtOH for genetic analyses.
Diagnosis. A medium-sized Brookesia species characterized by (1) body without a dorsal ridge (keel) and (2) presence of a complete series of laterovertebral pointed tubercles, with (3) the posteriormost laterovertebral pointed tubercle modified into a diamond-shaped pelvic shield, (4) nine pairs of laterovertebral pointed tubercles, (5) absence of enlarged pointed tubercles around the vent, (6) presence of poorly defined laterovertebral tubercles along the entire tail.
Brookesia brunoi differs from all other Brookesia species, except for Brookesia brygooi , B. decaryi , B. bonsi , B. valerieae , B. ambreensis , B. antakarana , B. griveaudi , and B. stumpffi by characters 1–3 as listed in the previous paragraph. Brookesia brunoi differs from B. valerieae by its smaller size (SVL up to 42.8 vs. 46–53 mm), a clearly defined pelvic shield (vs. poorly defined), and rounded supraocular cone projecting forward to the level of nostril (vs. pointed supraocular cone not projecting forward to the level of nostril). It differs from B. ambreensis by the number of laterovertebral pointed tubercles (9 vs. 11–12), presence of poorly defined laterovertebral tubercles on tail (vs. absence), clearly defined pelvic shield (vs. poorly defined); from B. antakarana by the number of laterovertebral pointed tubercles (9 vs. 12–13); and from B. griveaudi by its smaller size (TL up to 67.8 vs. 86–99 mm), presence of poorly defined laterovertebral tubercles on tail (vs. absence), and rounded supraocular cone (vs. pointed supraocular cone). Brookesia brunoi differs from B. stumpffi by its smaller size (TL up to 67.8 vs. 81–93 mm) and the presence of poorly defined laterovertebral tubercles on tail (vs. absence).
The most similar species to B. brunoi are Brookesia brygooi , B. decaryi and B. bonsi . Together with the morphologically highly distinct B. perarmata , these four species form a monophyletic group. Within this clade, B. brunoi can be distinguished by a combination of characters, especially by its lower number of laterovertebral pointed tubercles (9 vs. 10 in B. brygooi , B. decaryi , and B. bonsi ), absence of enlarged pointed tubercles around the vent (present in the other three species), the configuration of its cephalic crest, the lateral tail tubercles (poorly defined), and hemipenial morphology. A summary of these differences is provided in Table 2 View TABLE 2 .
Description of the holotype. Well preserved male with everted hemipenis and extruded tongue. Snout-vent length 40.3 mm; tail length 25.9 mm. Head with lateral, orbital and posterior ridges that form a cephalic crest; crest divided into three plains by a pair of longitudinal parasagittal ridges that start above the eyes converging at the posterior part of the crest resulting in a median plain (MP) on the top of the crest, surrounded by two triangular parietal plains (PP) on each side (see Fig. 1 View FIGURE 1 , C). Four pointed tubercles on each side of the posterior part of the crest, one at termination point of lateral ridge, one at termination point of parasagittal ridge, and two between the parasagittal and lateral ridges; two pairs of pointed tubercles on lateral surface of head: one supralabial pair, just above posterior angle of mouth, composed of two adjacent conical scales, the posteriormost the biggest, and one infralabial pair of tubercules below posterior angle of mouth. Each tubercle is composed by a single conical scale; orbital crest denticulated; two small and poorly marked inter-orbital ridges converging anteriorly, forming a “Vshaped” pattern; supra-ocular cone rounded and projects forward to level of nostril; supra-nasal cone does not reach as far forward as snout tip.
Dorsal surface of body flat (without a dorsal ridge or keel); 9 pairs of laterovertebral pointed tubercles form a complete longitudinal line on body; posterior-most (9th) enlarged pair of pointed laterovertebral tubercles project posteriorly and laterally to form a diamond-shaped pelvic shield above insertion point of hind limbs; l st -8th anterior pointed tubercles equally spaced and almost equal in size, perpendicular to body; no ventrolateral pointed tubercle just posterior to the vent at the tail base; reduced laterovertebral pointed tubercles on the tail.
Hemipenis short and stout, in general shape similar to that of B. brygooi as described by Brygoo & Domergue (1971) under the name B. ebenaui . Because only a single male is available and it is uncertain whether the organs are fully everted and turgid, the exact shape of the hemipenis is difficult to discern. No clear differentiation between truncus and apex. The sulcus spermaticus is short and deep, surrounded by a bulging basal lobe ("bourrelet basal" in Brygoo & Domergue 1971) and the lobular truncus. No field of denticulated papillae as in B. brygooi (Brygoo & Domergue 1971) is recognizable. A distinct denticulated rotula ("auricule bi-crêté of Brygoo & Domergue 1971; "denticulated lobe" of Raxworthy & Nussbaum 1995) is visible, much larger and more prominent than in B. brygooi where this rotula is usually smaller, less prominent and less strongly denticulated in the specimens we examined. In comparison to the hemipenes of B. brunoi and B. brygooi , the organ of B. decaryi (examined in ZSM 558/2001 and 560/2001) differs by a sulcus spermaticus that is continuous to the apex, absence of distinct rotulae or papillary fields, and presence of one very characteristic 1 mm long papilla-like protrusion of the apex, along with two much smaller such apical structures.
Coloration. According to the photographs ( Fig. 1 View FIGURE 1 ), the background coloration in life of the holotype specimen is beige, with exception of the flanks, the lateral side of the head, the ocular region, the throat and an indistinctly edged vertebral line that are darker. Several dark marks are present on the body: a median transversal black bar joins the summit of each supraocular cone, a median black mark extending from the inferior side of the posterior crest to the inferior side of the first pair of laterovertebral pointed tubercle, a pair of hourglass-shaped black marks running above the parietal ridge of the crest, one pair of black dots between the third and the fourth pair of laterovertebral pointed tubercles, one pair of black dots between the fifth and the sixth pair of laterovertebral pointed tubercle, large black dots below the eighth and the ninth pair of laterovertebral pointed tubercle (see Fig. 1 View FIGURE 1 , B), and a dozen tiny black dots on the flanks. The iris is faded orange with a thin central whitish ring bordering the pupil. Six months after fixation, the coloration was relatively similar to that in life, with the exception of the flanks that were darker, the dorsal side of the head, which was lighter, and the absence of tiny black dots on the flanks.
Variation. Based on the female paratype ZSM 889/2010: SVL 42.8 mm; tail length 24.9 mm + 1 mm (cut for molecular analyses). The color of the paratype after one year and a half in alcohol was similar to that at the time of collection. Ground color of head, body, tail and dorsal parts of limbs dorsally and laterally beige or brown, often becoming darker dorsally. The ground color is contrasting, with a mosaic of whitish, beige, and brown areas, which can be interpreted as leaf mimesis (or more in general as a mimesis to plant debris of the dry forest floor including also sticks and dried bark) ( Fig. 3 View FIGURE 3 ). The blotches on the back break up the background color providing discontinuity thus enhancing the overall cryptic appearance. No measurements are available for the female paratype specimen in the UADBA collection (ZCMV 12783).
Distribution, conservation and IUCN Red List status. The new species is currently known only from the type locality within the Anja Reserve (see Fig. 1 View FIGURE 1 in Crottini et al. 2011), although a recent Brookesia record from Ambovombé in the far south ( Hofmann 2012), ca. 375 km air distance from the type locality, seems to refer to B. brunoi . Furthermore, it is possible that some literature records of B. decaryi , B. brygooi , B. ebenaui and B. stumpffi from the central areas of Madagascar might actually refer to B. brunoi indicating that further investigations are required to better understand its actual distribution.
In the Ambalavao area we did not observe any mineral or precious stone extractions or collecting for the pettrade, but deforestation for agriculture, logging and cattle grazing is widely afflicting the area. Although relatively abundant in the Anja reserve (several individuals were observed in a few hours of active searching) it is possible that B. brunoi will qualify for inclusion in one of the threatened categories, depending on the actual size of its distribution area. However, due to the limited knowledge on this species we suggest to consider its conservation status as “Data Deficient” according to IUCN criteria ( IUCN 2001).
Habitat and habits. The holotype of B. brunoi and the UADBA paratype (ZCMV 12783) were found around 21:00 at a roosting height of about 30 cm from the leaf litter on small bushes in the forest of Anja Reserve. With the exception of the paratype specimen ZSM 889/2010, found dead in the jaws of a female Furcifer oustaleti , all other individuals sampled for the molecular analyses were found between 13:00 and 14:00 in the leaf litter (ACZC 1938, ACZC 1981, ACZC 1924) ( Fig. 3 View FIGURE 3 ), in shaded spots inside the forest fragment with closed canopy. Another individual was found at night around 21:00 at a roosting height of about 40 cm on small herbal bushes (ACZC 1934). No other Brookesia species were found around the type locality. Although inhabiting the humid leaf litter this new species is one of the few Brookesia species adapted to the semi-arid deciduous forest of Madagascar. The other species found in arid habitats of the west are B. bonsi , B. exarmata , B. decaryi , B. perarmata , B. brygooi and B. stumpffi . Other reptiles found in the forest of the Anja Reserve during our two visits (December 2009, December 2010) were: Paragehyra sp. aff. petiti, Paroedura sp. aff. bastardi, Phelsuma gouldi , Thamnosophis lateralis , Furcifer lateralis , Furcifer oustaleti , Oplurus quadrimaculatus , Madagascarophis meridionalis , Trachylepis vato , and Hemidactylus frenatus .
Etymology. A. Crottini dedicates this new species to Bruno Grassi in recognition of his love and support during these years. The specific name is thus a patronym, but in addition recalls the cryptic brownish coloration of the newly described species in Italian language.
Mitochondrial variation, differentiation and phylogenetic relationships. The molecular data confirms the attribution of B. brunoi to the B. decaryi group ( Townsend et al. 2009). The analyzed specimens of B. brunoi are genetically very uniform and show an intraspecific uncorrected divergence of 0.3%, while the analyzed specimens of B. brygooi and B. decaryi (including the sample from Tsingy the Bemaraha) are more heterogeneous and show intraspecific uncorrected divergence of 4.8% and 7.7% respectively, in the ND2 gene fragment sequences. The genetic distance between B. brunoi and the three other species of the B. decaryi group (molecular data of B. bonsi are not available for this gene) ranges between 17.6% (comparison between B. brunoi and B. brygooi ) and 19.7% (comparison between B. brunoi and B. perarmata ), and the genetic distance between B. brunoi and B. decaryi is 19.1%. Among the species of the analyzed species group the smallest genetic distance is observed between B. brygooi and B. decaryi (16.2%) and the highest value between B. perarmata and B. decaryi (20.8%). More details are provided in table 1.
the pairwise distance calculation for B. perarmata , B. brunoi , B. brygooi and B. decaryi . nc: not calculated. The phylogenetic analyses resulted in a tree with largely unresolved basal relationships ( Fig. 4 View FIGURE 4. 50 ) but with good support for the monophyly of the samples of the B. decaryi group, and for each species of this species group, hence clearly supporting the distinctness of B. brunoi from the other three analysed species. In the analysis of Townsend et al. (2009) B. bonsi was sister to B. decaryi based on DNA sequences of the ND4 gene available from the study of Raxworthy et al. (2002), and these two species were sister to B. brygooi . We could not include B. bonsi in our analysis due to the lack of samples and ND2 sequences for this species, but the combined evidence from this study and the tree of Townsend et al. (2009) suggests these species are related as follows: ( perarmata ( brunoi ( brygooi ( bonsi , decaryi )))).
In our analysis, samples of B. brygooi are organised in two mitochondrial lineages. Samples from Ankarafantsika belong to one lineage, while the samples of B. brygooi from Isalo and Kirindy belong to a second lineage, with an uncorrected average inter-lineage divergence of 5.9%. Furthermore, our analysis includes one sample from the Tsingy de Bemaraha that in our tree is sister to B. decaryi from the type locality Ankarafantsika, albeit with a high average divergence (11.3%). Because our sampling does not include B. bonsi , the sister species of B. decaryi ( Townsend et al. 2009) , we cannot decide whether the Tsingy de Bemaraha sample is to be assigned to B. decaryi , to B. bonsi , or possibly to a third, undescribed species. Neither B. bonsi nor B. decaryi have so far been reported from the Tsingy de Bemaraha, and a more detailed molecular and morphological study of this population is therefore necessary.
bonsi . The table also includes B. stumpffi which is the only other superficially similar Brookesia occurring in parts of western
Madagascar. Data for B. bonsi were taken from Ramanantsoa (1979).
B. brunoi B. brygooi B. decaryi B. bonsi B. stumpffi The description of B. brunoi sp. nov. adds a distinctive new species to the most basal subclade of the „typical“ Brookesia clade that contains B. decaryi , B. brygooi , B. bonsi and B. perarmata . All these four species inhabit dry deciduous forest areas of the west. On the contrary, B. brunoi is so far known to inhabit only the dry forest of the Anja Reserve, in the central high plateau of Madagascar, at an elevation of ca. 950 m a.s.l.. This small area (ca. 30 hectares) close to the town of Ambalavao has been protected since 1999, and is managed by the local community. Recent herpetological surveys in this area found evidence for the existence of several local endemics ( Crottini et al. 2011) and the present study represents a step further towards the knowledge and description of its endemic fauna.
Based on its morphological characteristics, B. brunoi in addition to its peculiar distribution, has at least two morphological characters that seem to be unique when compared with the other Brookesia species of the B. decaryi group: 1) the number of the laterovertebral pointed tubercles; and 2) the absence of enlarged pointed tubercles around the vent.
The molecular analyses suggest that this species group forms a well-supported monophyletic lineage with high genetic divergences observed between all four species (more than 16% uncorrected pairwise sequence divergence in the ND2), indicating a long divergent evolutionary history. It has been suspected that the genetic variability observed between the different populations of B. brygooi across the island reflects the existence of a species complex ( Townsend et al. 2009). However, the comparatively high genetic variability within B. brygooi , coupled with the lack of obvious morphological differences among the individuals analyzed in this study (see Appendix) suggests that this divergence might more appropriately be considered as intra-specific variability.
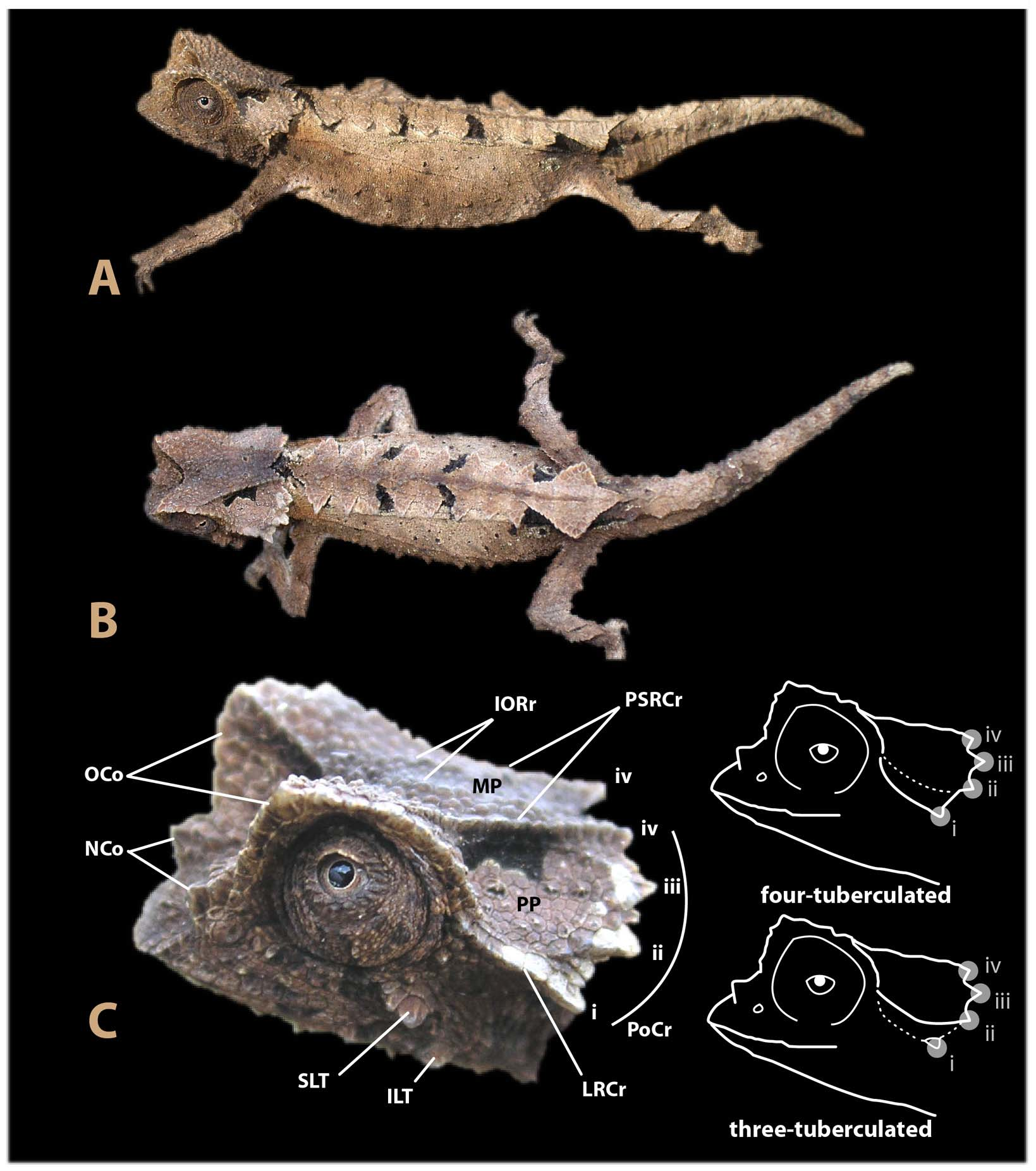


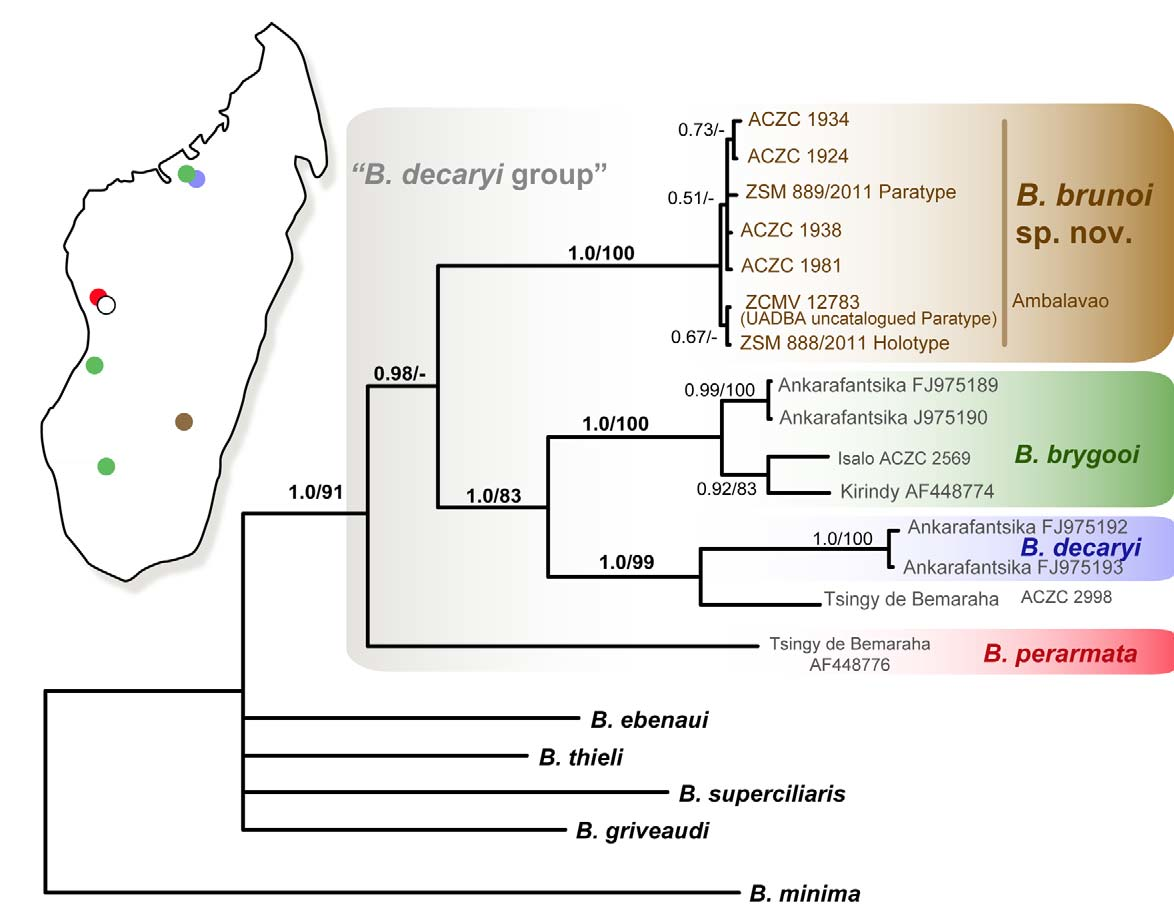
TABLE 2. Diagnostic character states of B. brunoi compared to those of its closest relatives B. brygooi, B. decaryi, and B.
| Laterovertebral pointed tubercles | 9 | 10 | 10 | 10 | 9–10 |
|---|---|---|---|---|---|
| Enlarged pointed tubercles around the vent | absent | present | present | present | absent |
| Laterovertebral tubercles along the entire tail | usually present but poorly defined | often only up to 2 tubercles anteriorly | typically distinct and strongly expressed on anterior two thirds | as far as known present along almost entire tail length | absent |
| Posterior part of the cephalic crest | four-tuberculated | typically three- tuberculated | four-tuberculated | possibly three- tuberculated | three- tuberculated |
| Hemipenis | Short and stout. Large and prominent denticulated rotula. No apical field of denticulate papillae. No apical projection. | Short and stout. Smaller and usually less prominent rotula, sometimes only weakly denticulate. Often with apical field of denticulate papillae. No apical projection. | Short and stout. No rotula. No field of papillae. Distinct and long papilla-like apical projection. | Rather elongated. Small slightly denticulate rotula. No field of papillae. Distinct but short papilla-like apical projection. | Elongated. Large central denticulate rotula. Two lateral lobes with denticulate papillae. No apical projection. |
| SVL [mm] | 40.3–42.8 | 42–52 | 43–53 | 37–40 | 46–55 |
| TL [mm] | 66.2–67.7 | 68–81 | 63–80 | 59–67 | 81–93 |
| Discussion |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.