
Blommersia variabilis, Pabijan, Maciej, Gehring, Philip-Sebastian, Köhler, Jörn, Glaw, Frank & Vences, Miguel, 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.207732 |
|
DOI |
https://doi.org/10.5281/zenodo.6189887 |
|
persistent identifier |
https://treatment.plazi.org/id/03F687F7-FF9F-6C1A-DAFD-9BA96EF1B40B |
|
treatment provided by |
Plazi |
|
scientific name |
Blommersia variabilis |
| status |
sp. nov. |
Blommersia variabilis View in CoL sp. nov.
Holotype. ZSM 237/2010 (field number FGZC 4305), adult male ( Fig. 5 View FIGURE 5 ), from Ambodivoahangy, northwest of Maroantsetra, northeastern Madagascar (coordinates: 15°17’23.8'' S, 43°37’13.0'' E, below 50 m a.s.l.), collected on 3 April 2010 by P.-S. Gehring, F. Glaw, J. Köhler and M. Pabijan.
Paratypes. Eleven adult males: ZSM 236/2010 (field number FGZC 4289), UADBA uncatalogued ( FGZC 4290), UADBA uncatalogued ( FGZC 4306), with same data as holotype; ZFMK 52608–52610, collected by F. Glaw and M. Vences on 18 March 1991 in the coastal town of Maroantsetra (15°26' S, 49°44' E, ca. 20 m a.s.l.); ZFMK 52612–52615, collected by F. Glaw and M. Vences on 19 March 1991 at a site slightly inland from the coastal village of Voloina (15°34' S, 49°37' E) which is located south of Maroantsetra; ZMA 19509 (field number FGMV 2002.2214) collected by M. Vences and A. Sarovy on 12 February 2003 in Maroantsetra.
Diagnosis. Assigned to the genus Blommersia in the Mantellidae by a combination of (1) presence of intercalary elements between ultimate and penultimate phalanges of fingers and toes (verified by external examination), (2) presence of femoral glands and absence of nuptial pads in males, (3) presence of a moderately distensible, inconspicuously coloured single subgular vocal sac in males, (4) small size (adult SVL <25 mm), (5) semiarboreal habits and calling behaviour from vegetation above stagnant water. Among species of Blommersia , B. variabilis is distinguished from B. blommersae by vomerine teeth present (vs. absent); from B. domerguei by different colouration (absence of light brown dorsum with dark longitudinal markings) and vomerine teeth present (vs. absent); from B. grandisonae by different colouration (absence of distinct black lateral line ventrally bordered by white, vs. presence) and vomerine teeth present (vs. absent); from B. kely by larger size (male SVL 19–25 mm vs. 14–16 mm) and vomerine teeth present (vs. absent); from B. sarotra by larger size (male SVL 19–25 mm vs. 15–17 mm) and vomerine teeth present (vs. absent); from B. angolafa by colouration (absence of uniformly brownish dorsal colour with white dots) and vomerine teeth present (vs. absent).
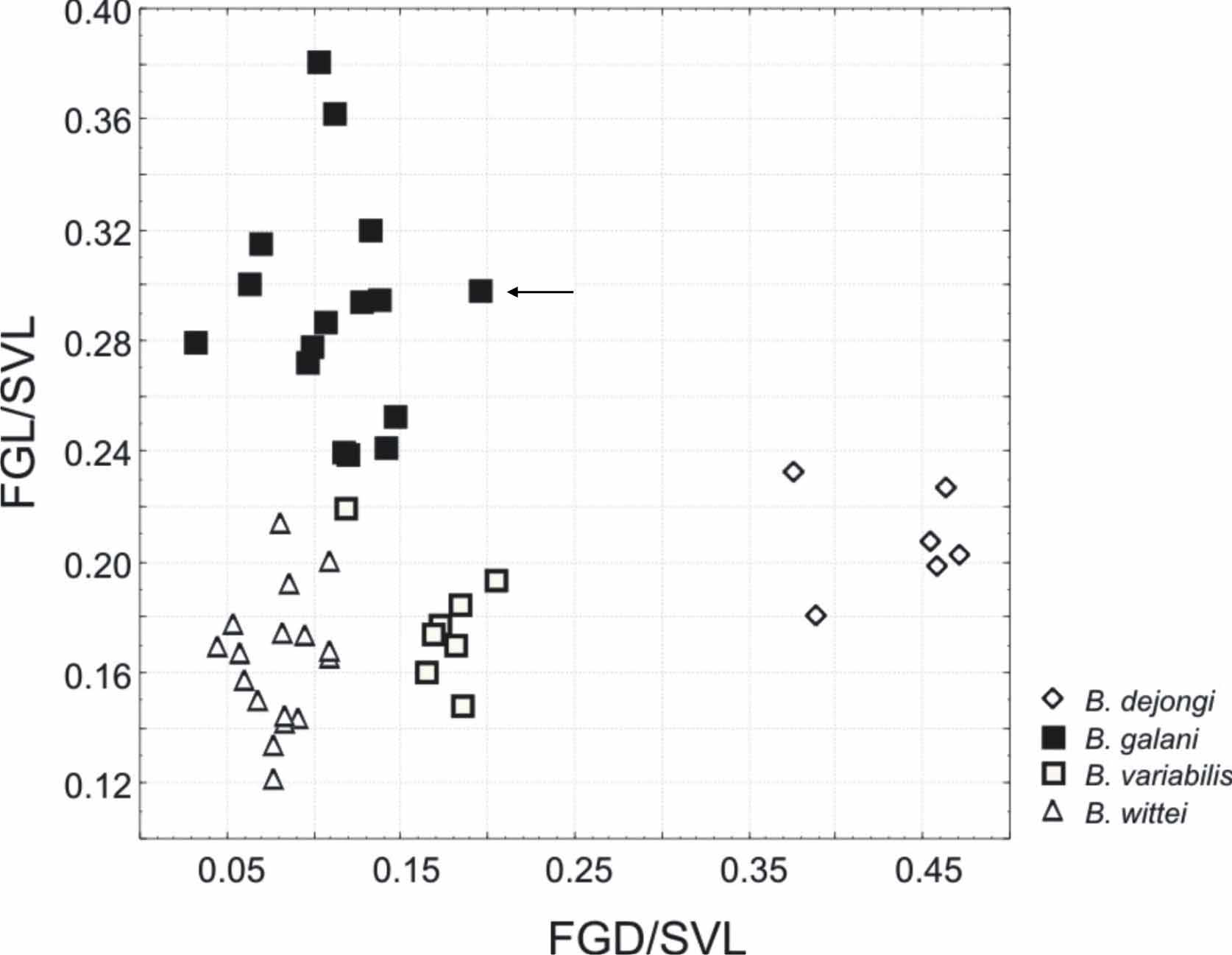
The new species is morphologically most similar to B. wittei from which it mainly differs by the wider distance between inner margins of femoral glands (FGD 2.8–4.5 mm vs. 1.6–2.0 in the type series of B. wittei ; see Vences et al. 2010; FGD 12–20% of SVL vs. 7–9%), connected metatarsalia in most specimens (vs. always separated by webbing), and advertisement calls consisting of a short series of 3-6 notes of a mean duration of 107 ms and a mean inter-note interval of 27 ms (vs. longer series of up to 25 notes with a mean duration of 23-46 ms and a mean interval duration of 30–72 ms). The new species forms a monophyletic group with the recently described B. dejongi and B. galani . It differs from B. galani by the presence of vomerine teeth (vs. absence) and smaller femoral glands (see Fig. 4 View FIGURE 4 ; femoral gland length 3.5– 5.2 vs. 4.8–9.2 mm). It differs from B. dejongi by a shorter distance between the inner margins of femoral glands ( Fig. 4 View FIGURE 4 ; FGD 2.8–4.5 mm vs. 7.8–9.8 mm). Furthermore, the new species differs from all nominal species of Blommersia by a substantial genetic divergence and by its advertisement call (as described below).
Description of the holotype. Specimen in a good state of preservation, tongue removed as tissue sample for molecular analysis. SVL 23.7 mm, for further measurements see Table 1. Body slender; head longer than wide, wider than body; snout rounded in dorsal and lateral views, nostrils directed laterally, slightly protuberant, nearer to tip of snout than to eye; canthus rostralis distinct, straight; loreal region slightly concave; tympanum distinct, rounded, its diameter 54% of eye diameter; supratympanic fold distinct posterior to tympanum, indistinct between eye and tympanum, curved; tongue removed, its shape therefore not verifiable; vomerine teeth present as two distinct protuberances immediately posterior-medially of choanae, maxillary teeth present; choanae small, rounded. Arms slender, subarticular tubercles single; fingers without webbing; relative length of fingers 1<2<4<3 on the right hand, 1<2=4<3 on the left hand; finger discs distinctly enlarged; nuptial pads absent. Hind limbs slender; tibiotarsal articulation reaches nostril when the hind limb is adpressed along the body; lateral metatarsalia largely separated; inner and outer metatarsal tubercles distinct; webbing formula (according to Blommers-Schlösser 1979) between toes 1(0.25), 2i (1.25), 2e(0.5), 3i (1.5), 3e(1), 4i (2.5), 4e(2.5), 5(0.75); relative length of toes 1<2<5<3<4. Skin on the upper surface smooth, without folds or ridges. No distinct enlarged tubercles in the cloacal region; ventral skin smooth. Femoral glands distinct, measuring 5.2 x 1.5 mm, of type 2 sensu Glaw et al. (2000), consisting of 17 distinct granules as verified from internal view (after reflexing of the ventral skin on thigh), distance between femoral glands 2.8 mm.
Table 1. Measurements of the type series of Blommersia variabilis sp. nov. and comparative specimens of B. wittei (all males). ZSM 237/2010 is the holotype, all other B. variabilis
specimens are paratypes. For abbreviations, see Material and Methods. Relative hindlimb length (RHL) is coded as follows: when hindlimb is adpressed along body, tibiotarsal articulation
reaches (1) anterior eye corner, (2) eye center, (3) between eye and nostril, (4) nostril, (5) snout tip.
After nine months in preservative, the dorsum appears greyish-brown because of many densely spaced, single dark melanophores. A translucent pink colour covers the entire dorsum, but is more prominent between and around the eye sockets and the tympanum. A thin, single medio-dorsal whitish line stretches from the neck in a posterior direction until the cloaca. There is a moderately distinct colour border between the flanks and the lighter dorsum. The hind limbs are light brown with four distinct dark brown crossbands on the thighs, three indistinct crossbands on tibia; tarsus light brown. The arms show indistinct, irregular dark patterns. Posterior to the tympanum a lateral dark streak runs along the supratympanic fold and ends at the forelimb insertion. Ventrally, the holotype was whitish with single, small and irregular dark melanophores. Throat almost uniformly whitish. Femoral glands largely white, contrasting with the yellowish colour of the inner thighs.
In life the holotype ( Fig. 5 View FIGURE 5 ) had a brown dorsum with a reddish hue on the anterior half of the body, including the head and neck, whereas the posterior is grayish-brown. Indistinct, gray crossbands line the limbs and a distinct white vertebral stripe started in the neck region and continued down the length of the body. Iris golden dorsally, ventrally poorly recognizable due to the widely opened pupil. Ventrally, the throat is yellow, but the rest of the body is white with some yellow mottling along the sternum and along the sides. The undersides of the front limbs are pinkish-blue. The ventral side of the hindlimbs is a darker blue, especially on the digits and joints. The ventral side of the thighs is pinkish-blue with yellow, clearly discernible femoral glands. The skin on the venter was translucent such that larger blood vessels could be easily discerned. The ventral colouration extends slightly onto the flanks, merging rather indistinctly with the dorsal brown colour.
Remark. This species was referred to as Blommersia sp. aff. blommersae “Maroantsetra” by Glaw & Vences (2007), Blommersia sp. aff. wittei (Maroantsetra) by Vences et al. (2006), and Blommersia sp. 2 Maroantsetra by Vieites et al. (2009) and Vences et al. (2010).
Variation. Morphological variation of nine paratypes is provided in Table 1. In general the morphology of the paratypes was similar to the holotype. However, there is some variability in the separation of the lateral metatarsalia. For instance, in ZFMK 52608 they are separated, whereas in other specimens they are partly connected, and ZFMK 52612 shows connected metatarsalia on the left foot and nearly completely separated metatarsalia on the right foot. Dorsal colouration of the paratypes is rather variable. The reddish hue characterizing the holotype is not typically observed in other specimens which are usually light brown with dark brown markings. A narrow white median line can be present on the dorsum as well as a broader band that becomes triangular at the anterior part of the dorsum, covering the entire surface of the head. Brown or grey markings and spots can be present on the venter. In 2003 we also collected one specimen of very large body size in Maroantsetra, mixed with normal-sized specimens. Because of a labeling error we cannot ascertain with full reliability the current identity of this specimen and therefore did not include it in Table 1, almost certainly it is the male ZMA 19511 of 29.0 mm SVL with largely separated outer metatarsalia (intermediate between the connected and separated state).
Etymology. The species name variabilis is an adjective referring to the remarkable variation observed in various characters of this new species, especially in the separation or connection of the lateral metatarsalia which otherwise is considered to be a very stable character within species of mantellids.
Natural history. At the type locality in the vicinities of Ambodivoahangy village, B. variabilis was common outside of the forest in cultivated landscape. It was not seen or heard within primary or selectively logged forest. Calling males were observed in dense secondary vegetation on the borders of partially inundated ricefields, about 30 cm to 1 m off the ground. The male holotype was discovered while calling within a dense bush at the edge of a rice field. During the day specimens were difficult to find, hiding underneath the leaves of plants. Calling was most frequent in the evening and at night, but was also occasionally heard during the day, especially after rains. Calls were often intermingled with choruses of Aglyptodactylus sp., Guibemantis cf. kathrinae and Ptychadena mascareniensis . At Maroantsetra it is a common species living in ditches covered by dense vegetation within the town itself. Near Voloina we heard and collected the species in flooded areas in a mosaic of rice paddies, shrubs, and remains of primary rainforest. A single clutch of eggs deposited on the surface of a leaf was discovered within the village of Ambinanitelo on 1 April 2010. Mating in this species takes place on a leaf usually overhanging lentic water and has been observed by us in Voloina. As in other Blommersia , the male and female sit vertically on a leaf, the male on top of the female with his legs extending over the female’s dorsum ( Glaw & Vences 1994).
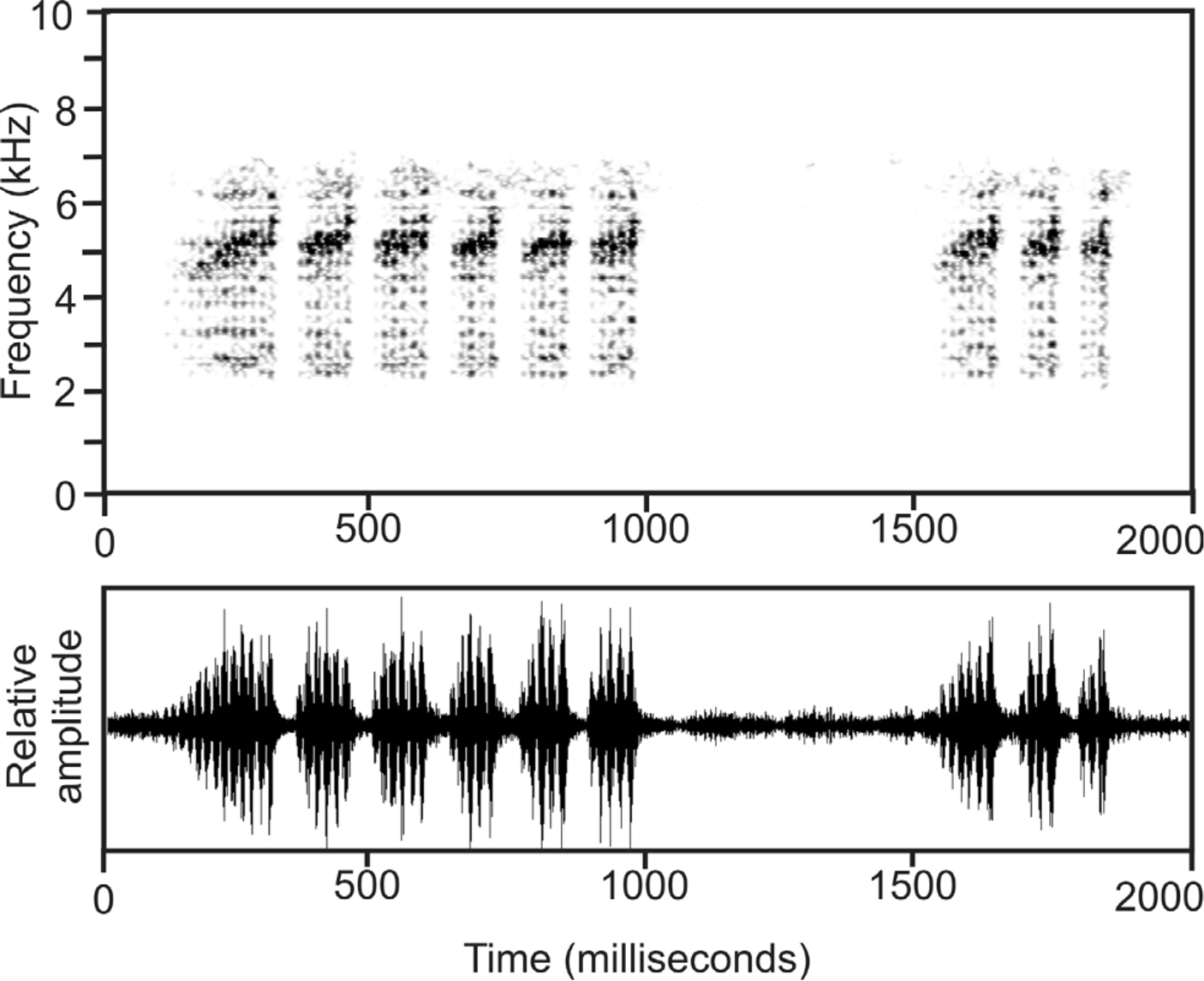
Vocalization. The advertisement call of B. variabilis ( Fig. 6 View FIGURE 6 ) was recorded at the type locality (Ambodivoahangy) on 3 April 2010 at an estimated air temperature of 26°C. It consists of a series of short inharmonious notes, repeated at regular intervals in fast succession with a repetition rate of approximately 7.4 notes/second. Each call is composed of 3–6 notes, whereas in most cases a long call is followed by a shorter call containing fewer notes, in combination resulting in a call series with two calls. Within these call series, the interval between first and second call ranges between 500 and 720 ms. Within calls, the initial note is longer and contains more pulses than subsequent secondary notes. Numerical call parameters are as follows: call duration 344–887 ms (532 ± 219; n = 9); note duration 54–223 ms (107 ± 48; n = 23); pulses/note 3–14 (6.4 ± 3.0; n = 23); inter-note interval 21–33 ms (26.7 ± 3.2 ms; n = 19). Overall frequency is distributed in a broad band from app. 2000–6500 Hz, with a dominant frequency at around 5100 Hz. Calls were repeated at irregular intervals. Similar calls were recorded in 1991 in Maroantsetra, but due to the poor quality of the recordings we refrain from a detailed analysis.
Detailed call data for various populations of B. galani , B. dejongi , and B. wittei are presented in Vences et al. (2010). In comparison, the call of B. variabilis differs from that of B. wittei by longer note duration, shorter internote intervals and a lower note repetition rate. Calls of B. galani differ from those of B. variabilis by shorter notes and much longer inter-note intervals. In calls of B. dejongi , notes are barely spaced and partly fused and separate pulses within notes are unrecognizable.
Distribution. B. variabilis is known from (1) Maroantsetra, (2) Voloina, (3) Ambodivoahangy and (4) Ambohinantely (15°20'50.71'' S, 49°35'02.84'' E). The largest straight line distance between any of these sites is approximately 30 km (Ambodivoahangy-Voloina). The altitude was not precisely measured at the collecting localities, but all except Voloina are situated below 50 m a.s.l. We estimate an elevation of 100-300 m a.s.l. for the collecting locality near Voloina. The Voloina site lies close to a medium-sized river, but due to the lack of reliable coordinates for our precise collecting locality in 1991, we cannot ascertain if our samples originate from north or south of the river. The remaining three sites are in the immediate vicinity of the Antainambalana river.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |