
Bennelongia timmsi, Martens & Halse & Schön, 2013
|
publication ID |
https://doi.org/ 10.5852/ejt.2013.66 |
|
publication LSID |
lsid:zoobank.org:pub:5270B024-84C1-4CD5-BC18-4AF37D2E1045 |
|
DOI |
https://doi.org/10.5281/zenodo.3846924 |
|
persistent identifier |
https://treatment.plazi.org/id/4F6A6E8F-5636-4290-85A4-B234D5DA4466 |
|
taxon LSID |
lsid:zoobank.org:act:4F6A6E8F-5636-4290-85A4-B234D5DA4466 |
|
treatment provided by |
Carolina |
|
scientific name |
Bennelongia timmsi |
| status |
sp. nov. |
Bennelongia timmsi sp. nov.
Figs 4-11 View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig
urn:lsid:zoobank.org:act:4F6A6E8F-5636-4290-85A4-B234D5DA4466
Diagnosis
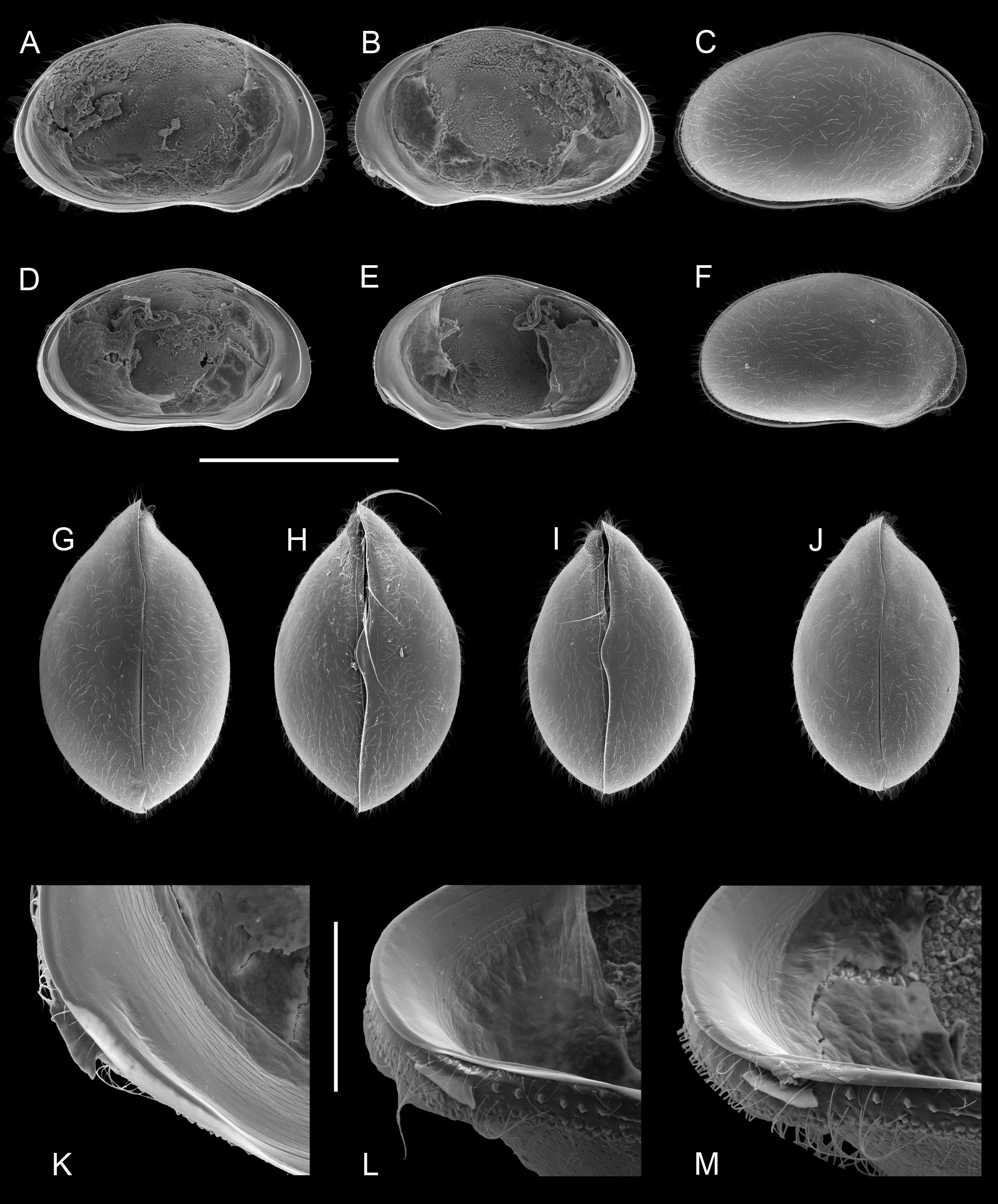
Valves in inner view ( Fig. 4 View Fig A-B, D-E) relatively high, with greatest height situated well in front of the middle; ventral margin anteriorly with well-pronounced mandibular curve. LV ( Fig. 4A, D View Fig ) with anterior il not overlapping. RV ( Fig. 4B, E View Fig ) with antero-ventral lapel subtriangular, asymmetrically produced with a ventral point ( Fig. 4 View Fig K-M). Carapace in dorsal and ventral views ( Fig. 4 View Fig G-J) with greatest width in the middle, hirsute, anteriorly with a mild rostrum; in lateral views ( Fig. 4C, F View Fig ) showing a clear anterior LV>RV overlap.
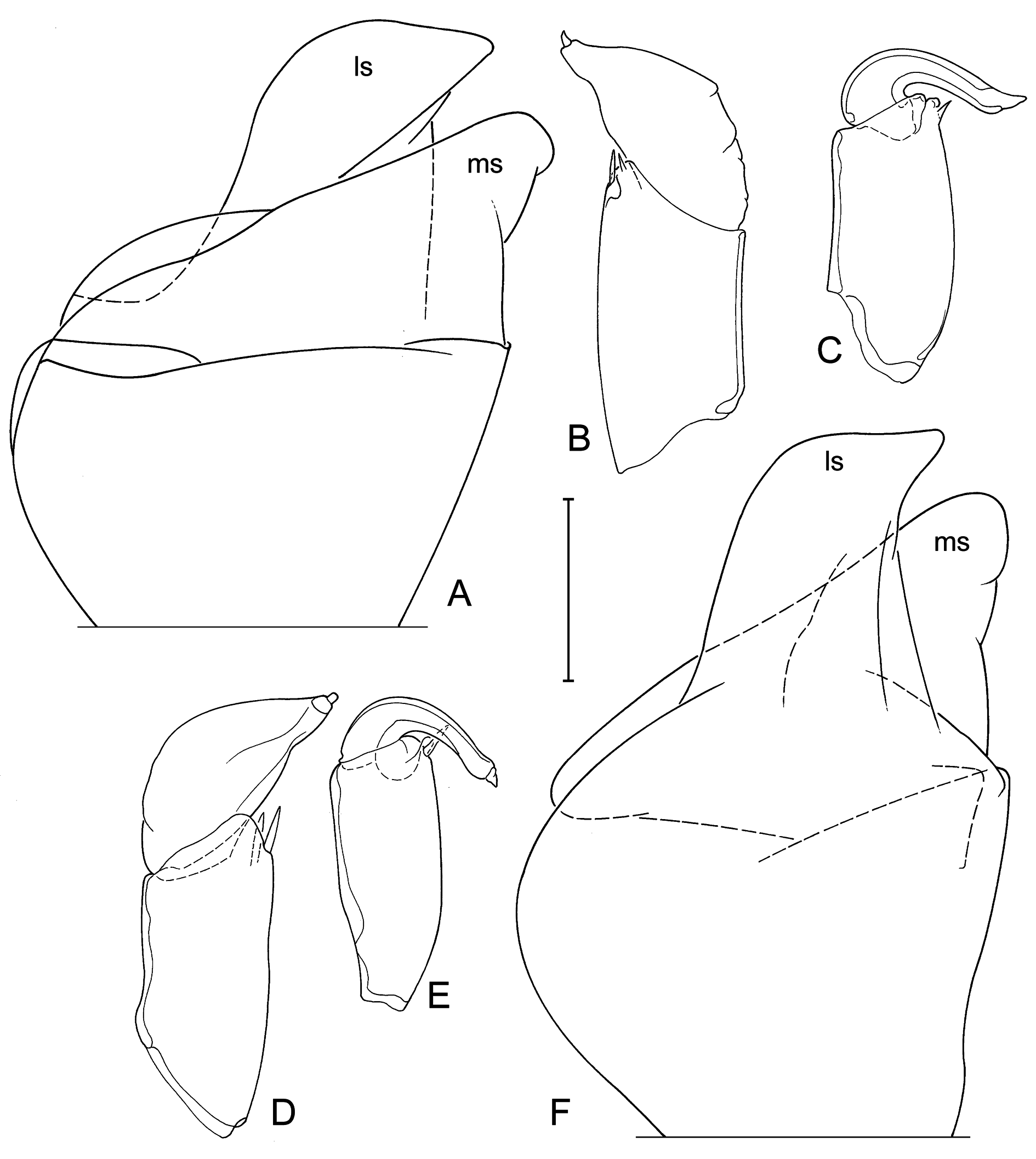
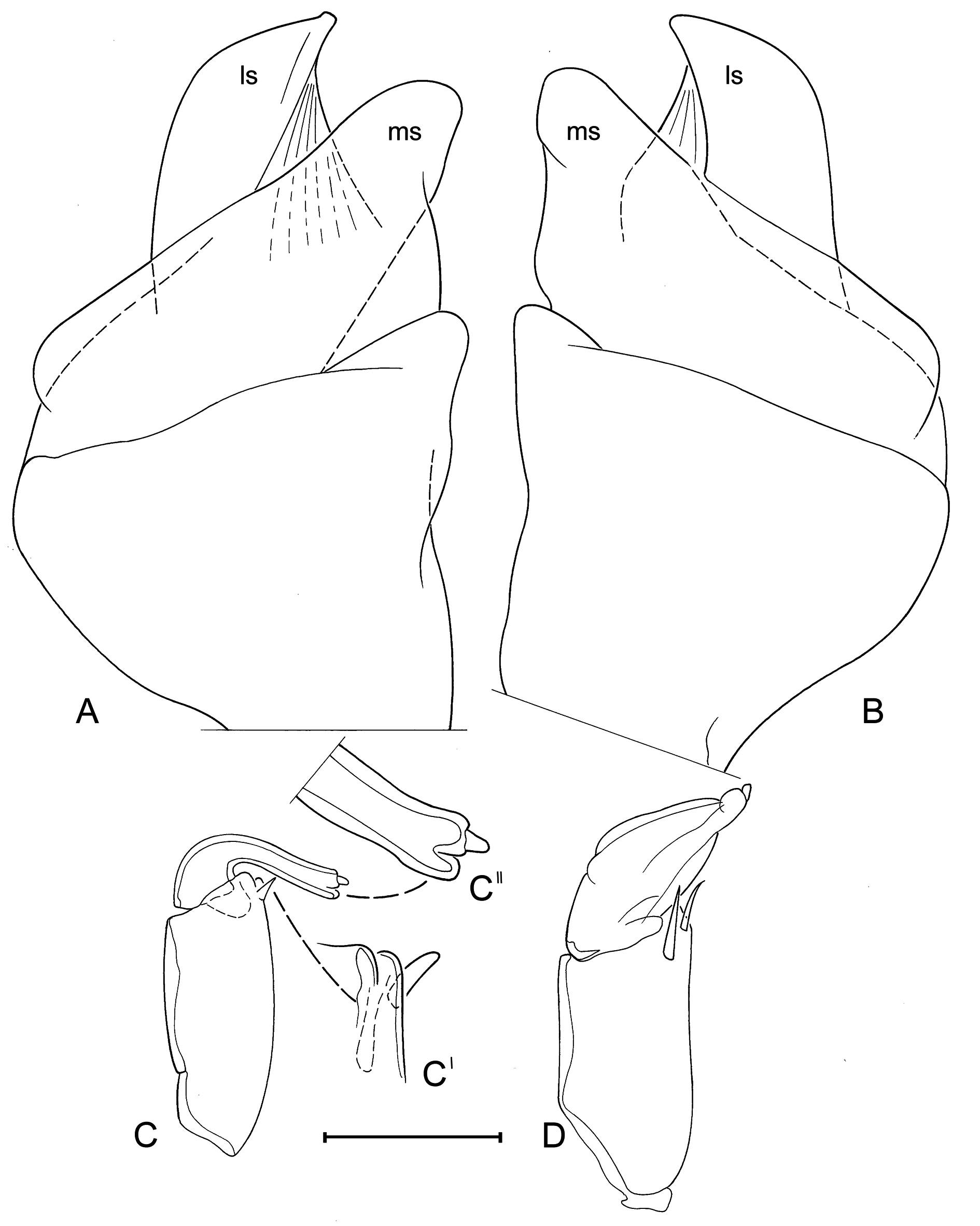
Hemipenes (holotype: Fig. 8F View Fig ) mostly symmetrical, with ls protruding well beyond ventral tip of ms, ls with broad base, ventrally bluntly beak-shaped. Right prehensile palp (holotype: Fig. 8D View Fig ) with distal segment elongated, with dorsal margin evenly rounded. Left prehensile palp (holotype: Fig. 8E View Fig ) with distal segment elongated, reaching beyond ventro-apical margin of proximal segment with at least half of its length.
Etymology
This species is named after Prof. Brian V. Timms (Newcastle, Australia), in recognition of his vast contribution to the knowledge of Australian non-marine crustaceans in general, and of phyllopods from temporary pools in particular. Prof. Timms also collected the material of the present species from a series of pools on various rocky outcrops in WA.
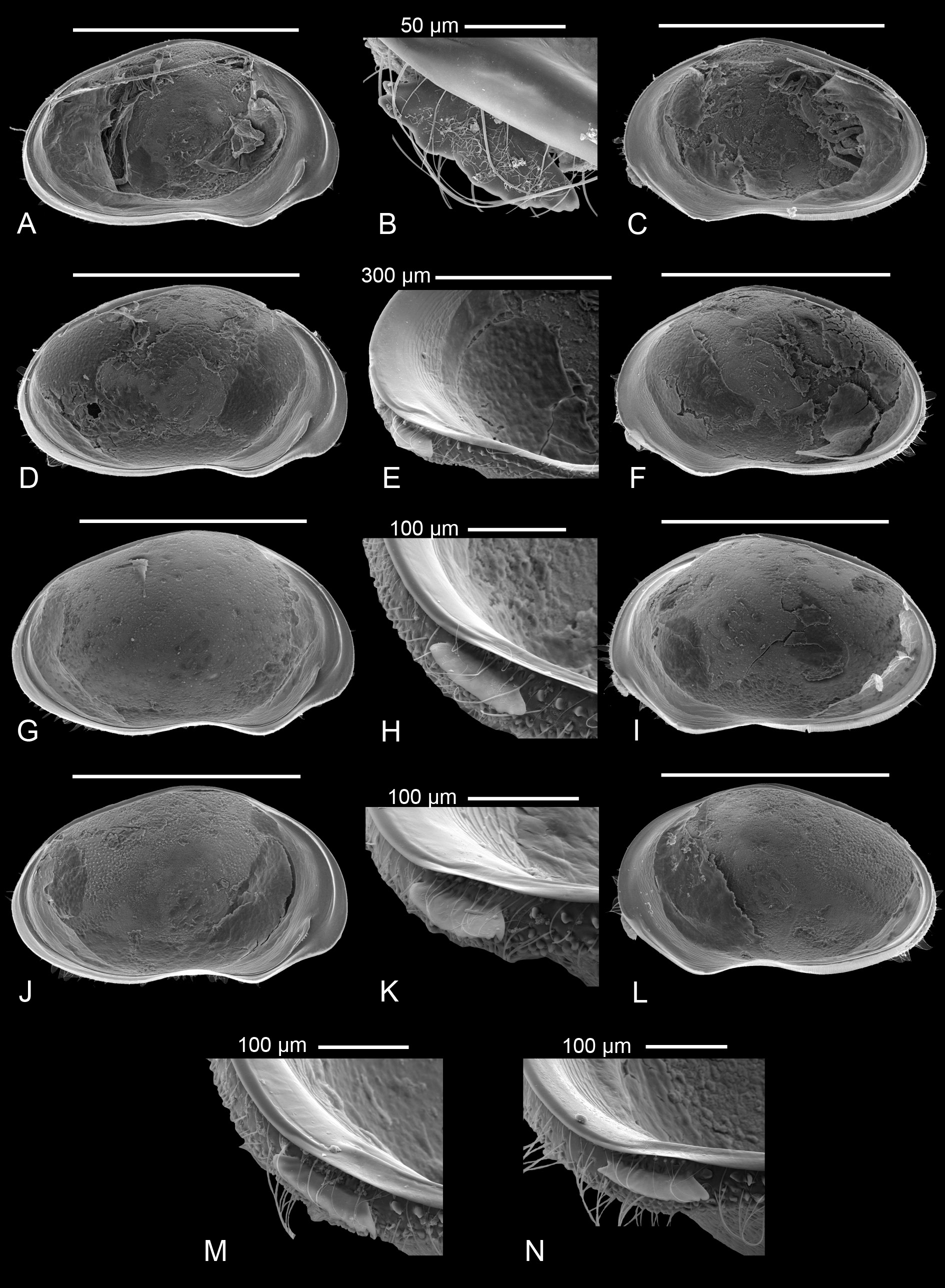
Measurements (all measurements in µm – see Table 1 View Table 1 for measurements of all specimens illustrated with SEM)
Holotype ♂ ( WAMC52228 ): RV: L = 1318, H = 783. LV: L = 1378, H = 817.
Allotype ♀ ( WAMC52229 ): RV: L = 1510, H = 913. LV: L = 1600, H = 965.
Type locality
Rock pools on Wave Rock, WA, ca. 2 km E of Hyden. Approximate coordinates: 32º 27’S 118º 54’ E (WGS 84). Material handpicked from pools by B.V. Timms on 23 Jul. 2010 (sample BVT/10/05).
Type material
Holotype
♂ ( WAMC52228 ), with soft parts dissected in a sealed slide and valves stored dry in a micropalaeontological slide.
Allotype
♀ ( WAMC52229 ), with soft parts dissected in a sealed slide, and valves stored dry in a micropalaeontological slide.
Paratypes
Numerous males and females from the type locality, either dissected and stored as the holotype, or as carapaces used for SEM ( WAMC 52230-52237, OC.3312-3316). Ca. 60 ♂♂ and ♀♀ in EtOH as bulk paratypes ( WAMC 52238).
Other material investigated
All material from WA, collected by B. V. Timms.
Paynes Find Rocks. Approximate coordinates: 29º10’ S, 117º40’ E (sample BVT /10/01), collected by B. V. Timms on 23 Jul. 2010 (one ♀ - WAMC 52239).
Grahams Rock. 32º28’ S, 119º03’ E (sample BVT /10/02), collected by B. V. Timms on 23 Jul. 2010 (six ♂♂ and ♀♀, WAMC 52240-52244; OC.3317).
Anderson Rock. 32º10’ S, 118º51’ E (sample BVT /10/03), collected by B. V. Timms on 23 Jul. 2010 (one ♂, WAMC 52245).
Burracopin Rock. 31º24’ S, 118º27’ E (sample BVT /10/04), collected by B. V. Timms on 26 Jul. 2010 (six ♂♂ and ♀♀, WAMC 52246-52249; OC.3318-3319).
King Rocks. 32º19’ S, 119º09’ E (sample BVT /10/06), collected by B. V. Timms on 23 Jul. 2010 (one ♂ and two ♀♀, WAMC 52250-52252).
Yorkrakine Rocks. 31º25’ S, 117º30’ E (sample BVT /10/07), collected by B. V. Timms on 27 Jul. 2010. Mt Madden Rock. 33º14’ 22” S, 119º50’ 33” E (sample BVT /10/08), collected by B. V. Timms on 01 Aug. 2010 (11 ♂♂ and ♀♀, WAMC 52253-52262; OC.332-3321).
Differential diagnosis
Bennelongia timmsi sp. nov. can be distinguished from most species of the B. barangaroo lineage by the triangular and ventrally pointed lapel on the RV and the strongly sinuous ventral valve margins. The lapel of B. scanloni sp. nov. is also subtriangular and ventrally protruding, but it is rounded, thus looking drop-shaped in internal (non-tilted) view.
Bennelongia timmsi sp. nov. can moreover be distinguished from B. gnamma sp. nov. by the less high and less rounded valves and by the ls of the hemipenes, which protrudes well beyond the ms (subequal in B. gnamma sp. nov.).
Additional notes on cryptic species
As was described above, five genetic clusters are recognised in this species (A1-5, Fig. 2 View Fig ). According to the calculations of the 4 theta rule, three cryptic species were found in B. timmsi sp. nov. with molecular methods, but no morphological diagnostic features could be found. Cryptic species A1 occurred in BVT/10/02, 03 and 07. Cryptic species A3 was found in BVT/10/02, 04, 06 and 08. Cryptic species A2+A4+A5 occurred in BVT/10/03 and 05 and is used here to characterize B. timmsi sp. nov. with BVT/10/05 (Wave Rock) as type locality. Note that BVT/10/02 (Grahams Rock) and BVT/10/03 (Anderson Rock) hold at least two sympatric clades/cryptic species each. In order to establish beyond reasonable doubt that the specimens belonging to these clusters and cryptic species are indeed morphologically indistinguishable, long series of specimens are illustrated.
Sample BVT/10/05 from pools on Wave Rock appeared to contain only one genetic cluster and cryptic species and, for this reason, Wave Rock was chosen as type locality. We then proceeded with two different approaches: (1) to dissect a series of males from this sample to test whether male reproductive organs (hemipenes, prehensile palps) showed uniformity within one cluster/cryptic species; (2) we checked for potential differences in the morphology of the valves of specimens belonging to different populations and/or shown to belong to different clusters/cryptic species.
Type specimens
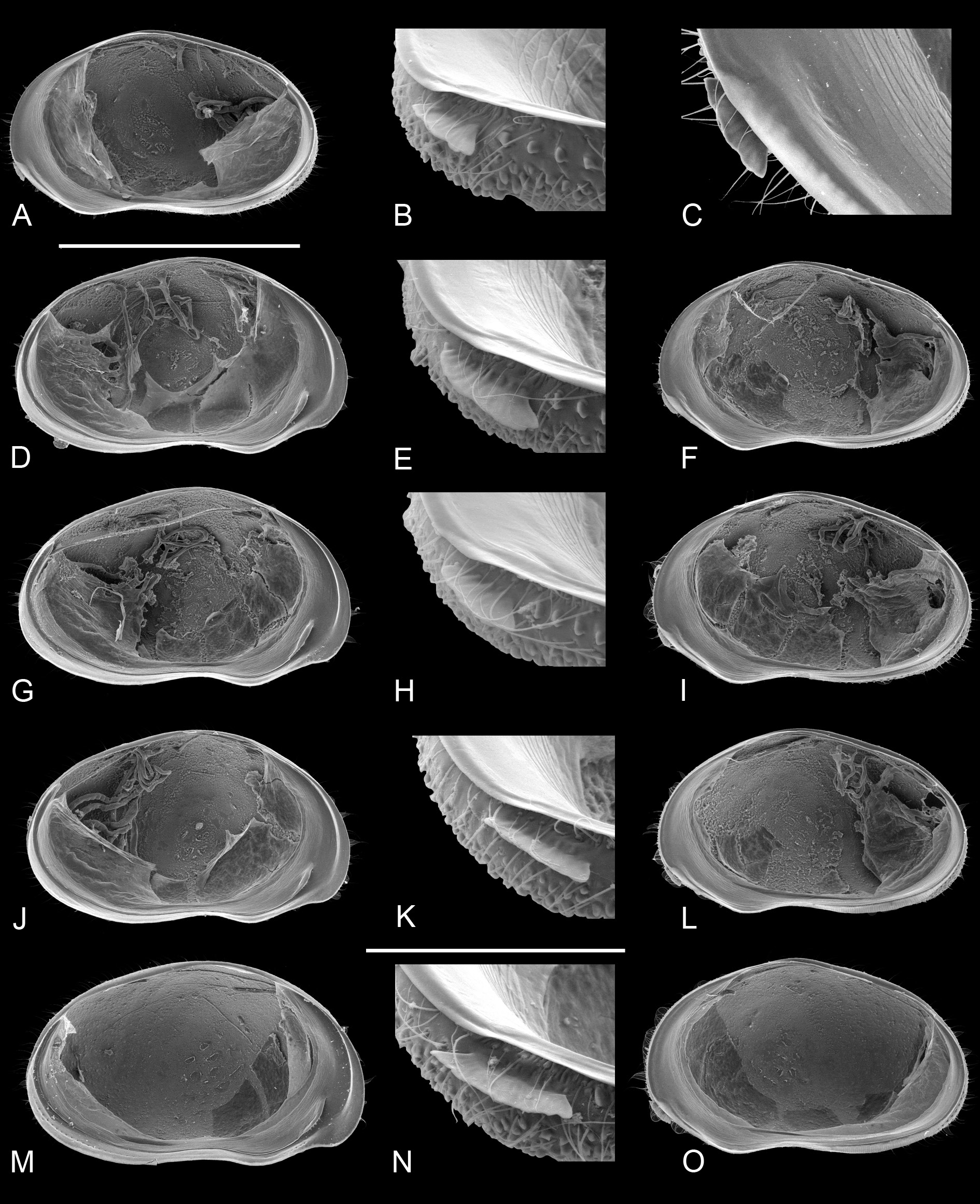
Valves and carapaces of males and females of the type population (in sample BVT/10/05) were illustrated ( Fig. 4 View Fig ) and this morphology defines the species. We then dissected several males from the same sample and population and illustrated the soft part and valve morphology. Shape of valves and size and shape of the antero-ventral lapel on the RV were most similar and indeed almost indistinguishable ( Fig. 5 View Fig ). In all male specimens the valves have the shape described in the diagnosis above. The lapels are all elongated subtriangular, with a more or less serrated distal margin. In tilted perspective some lapels appear to be shorter than others (e.g., the lapel in Fig. 5B View Fig appears shorter than in 5E), but this is almost entirely a matter of distorted perspective depending on how the valves were positioned when the photographs were taken (the same lapels appear almost equally long in non-tilted views - Fig. 5A and 5F View Fig , respectively).
However, there are significant differences in soft part morphology. Whereas the shapes of the hemipenisoutline and of the left prehensile palps are fairly uniform in the different specimens ( Figs 8A, C View Fig , E-F; 9A, C-D, F), the second segment of the right prehensile palps ranges from elongated sub-triangular with almost equally rounded distal margin ( Fig. 8B, D View Fig ), to sub-rectangular with a clear blunt corner in this margin ( Fig. 9B, E View Fig ). It is not clear to what extend these differences are a biological reality, or whether the differences are distortions of the limbs caused by different positions in the slides. The differences are sufficiently small to be accepted as part of intra-specific variability, yet future investigations should take this variability into account. The morphology in the holotype (WAMC52228 – right prehensile palp in Fig. 8D View Fig ) determines the specific morphology.
One male (WAMC52232 – Fig. 10 View Fig A-D) had an aberrant morphology, with the terminal segment of the right prehensile palp ( Fig. 10D View Fig ) being even more elongated and with especially the terminal segment of the left prehensile palp being distally bilobed ( Fig. 10C, C View Fig ”), a morphology never before encountered in Cyprididae . Nevertheless the valves of this male show no differences with other type specimens ( Fig. 5 View Fig A-C).
Morphology within different clades/cryptic species
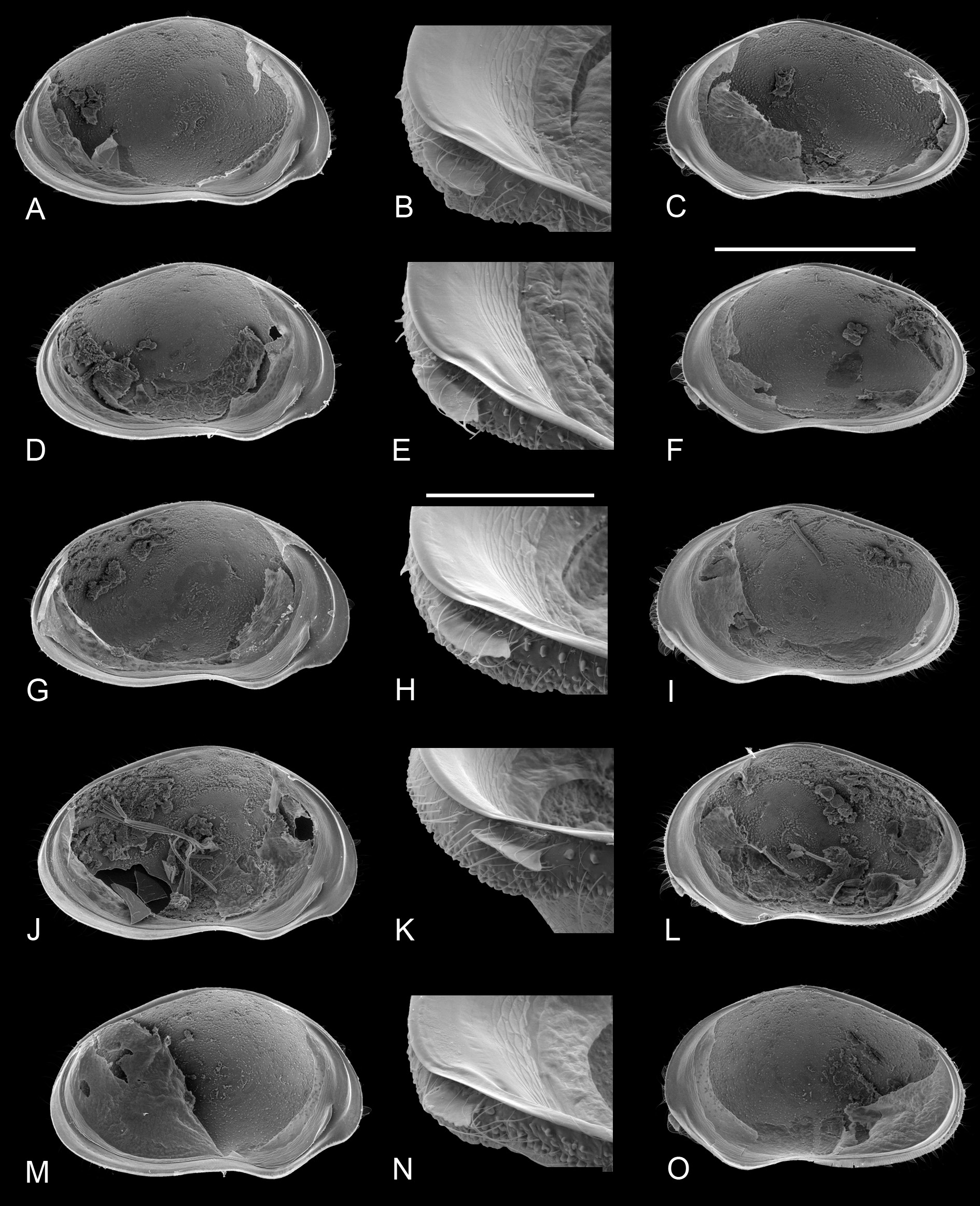
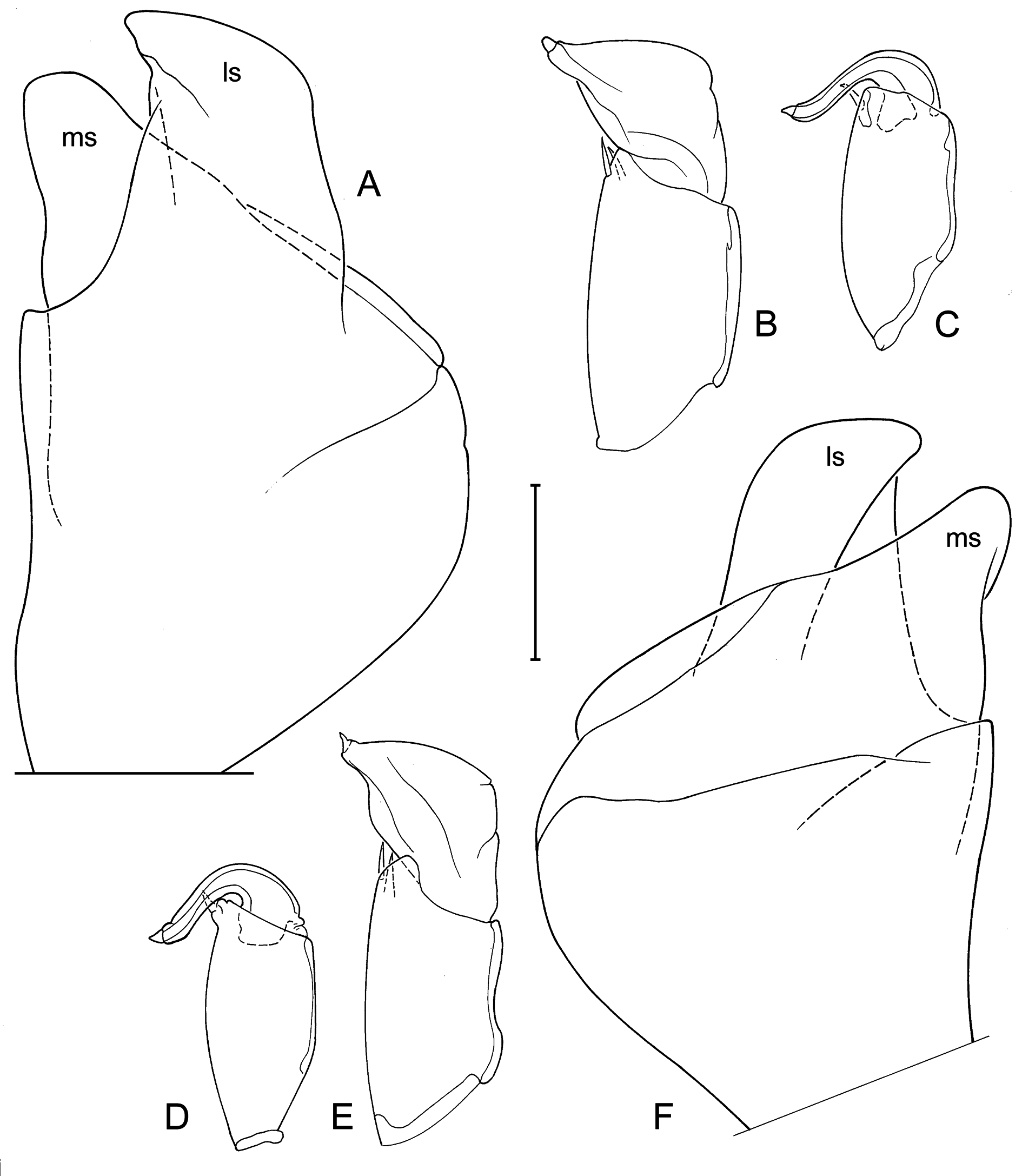
Valve morphology of specimens for which molecular clades are known (A1: Fig. 6 View Fig A-C; A2: Fig. 6 View Fig D- I; A3: Fig. 6 View Fig J-L; A5: Fig. 6 View Fig M-O, 7A-C) and for specimens from different populations for which no molecular data were available ( Fig. 7 View Fig D-N) again show no constant differences that could be used as identifying characters. There is some variability in size, shape and degree of crenulation of the lapel, but insufficiently so to use such features to characterise different clades/cryptic species.
One male specimen from BVT/10/02 and thus belonging to either cryptic species A1 or A3, had a right prehensile palp with a terminal segment clearly showing a blunt angle on the distal margin ( Fig. 11C View Fig ), while a male from BVT/10/06, and thus most likely belonging to clade A2, had a more elongated segment there with a more rounded distal margin ( Fig. 11G View Fig ) as in the holotype. In both of these specimens, the terminal segment of the left prehensile palp is slightly shorter than in the type specimens ( Fig. 11D, F View Fig ). Hemipenis outlines ( Fig. 11 View Fig A-B, E) are indistinguishable from those in the types.
Ecology and distribution
Bennelongia timmsi sp. nov. is a typical rock pool species and occurs in fresh water in gnammas on various rocky outcrops in the south/central part of western Australia. Although it appears to be limited to this restricted area, it seems to be quite common there.


| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |