
Baeolidia japonica Baba, 1933
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3802.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:4095CA09-8EA4-4941-8286-32E95F0206AE |
|
DOI |
https://doi.org/10.5281/zenodo.6134200 |
|
persistent identifier |
https://treatment.plazi.org/id/ED36FA51-A035-FF88-FF1B-F8DEE640F961 |
|
treatment provided by |
Plazi |
|
scientific name |
Baeolidia japonica Baba, 1933 |
| status |
|
Baeolidia japonica Baba, 1933 View in CoL
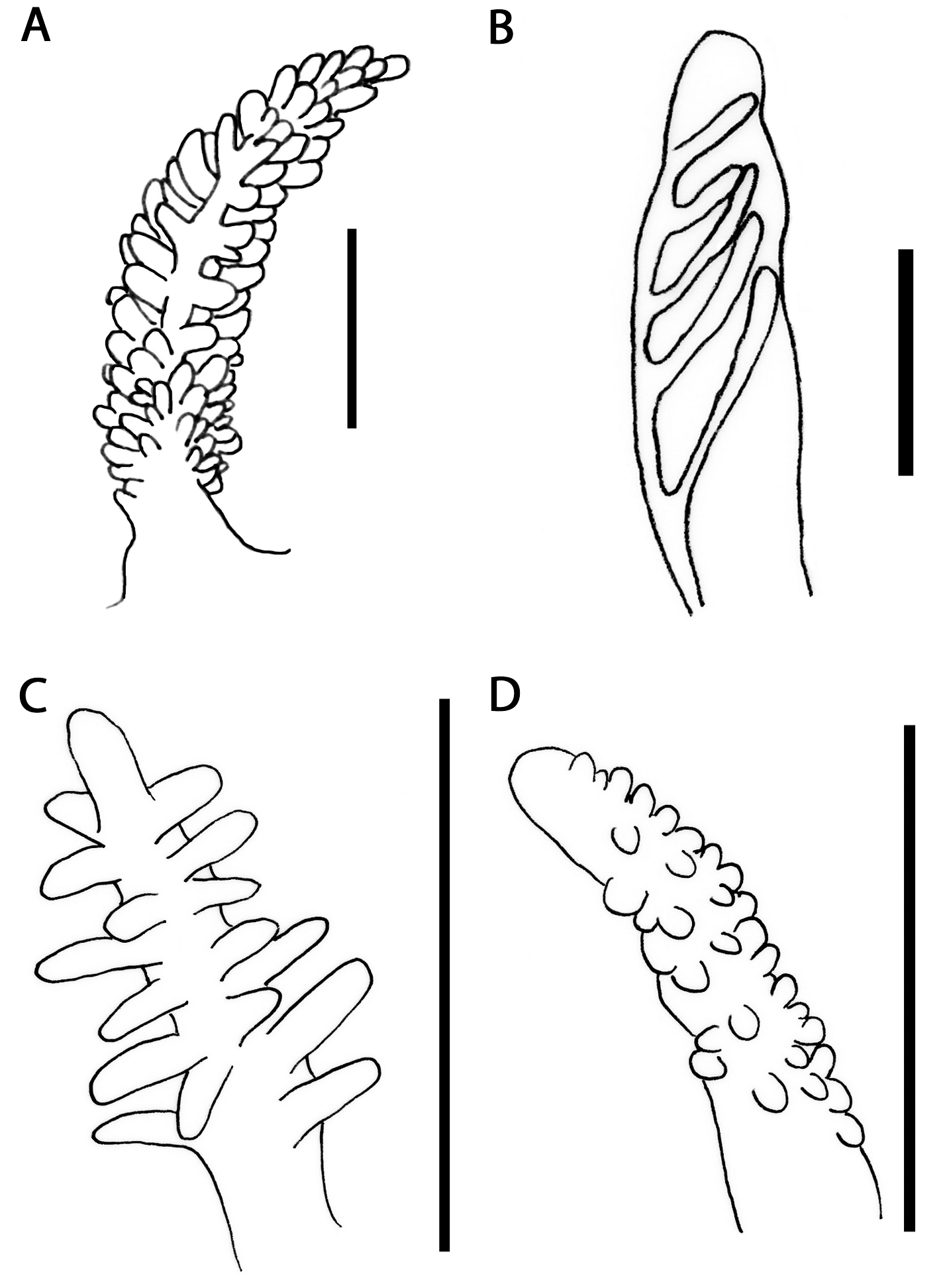
( Figs. 2 View FIGURE 2 D–E, 3C, 4E–F, 6C).
Baeolidia japonica Baba, 1933: 282 View in CoL , Fig. 8 View FIGURE 8 .
Spurilla japonica (Baba, 1933) View in CoL : Rudman 1982, 193. Berghia japonica (Baba, 1933) View in CoL : Gosliner 1985, 261.
Type locality. Tomioka Bay, Japan.
Type material. To our knowledge no type material remains. We designate the specimen CASIZ 1814520 from Ose, Japan, as the neotype in order to avoid confusion with similar species.
Material examined. Neotype: CASIZ 1814520, one specimen, dissected, 2 mm in length preserved, immature, Japan, Ose, collected by Akira Kawahara, 10 February 2010. Other material: CASIZ 186795, one specimen, dissected, 8 mm in length preserved, Marshall Is., collected by Scott Johnson, 12 September 2011; CASIZ 181357, one specimen, dissected, 5 mm in length preserved, Marshall Is., collected by Scott Johnson, 27 December 2008.
Geographical distribution. Originally described from Tomioka Bay, Japan (Baba 1933) it is also known from Shimoda ( Nakano 2004), the Ryukyu Is. ( Ono 2004), Indonesia (Gosliner et al. 2008), the Marshall Is. (present study), Philippines (Gosliner et al. 2008), and Papua New Guinea (Gosliner et al. 2008).
External morphology ( Figs. 2 View FIGURE 2 D–E, 3C): Body short, broad, tapering close to posterior end of foot. Foot corners tentaculiform. Body colour brown or translucent white. Bright white spots over dorsum, forming continuous patch from pericardial area to posterior end of foot. Tiny white spots form white mark over head. Rhinophores, oral tentacles and foot corners translucent white. Rhinophores shorter than oral tentacles. Rhinophores full of elongate, white papillae ( Fig. 3 View FIGURE 3 C); apex white. Oral tentacles with white spots. Cerata moderately short, flattened, almost leaf-like. Cerata usually brownish with white spots, or translucent covered by white pigment; density of white spots varies; some specimens with bluish band on anterior side; a minute white ring on apex. Cerata in two arches followed by rows, leaving a distinct gap between pre and post-pericardial groups. Each group with 4–11 cerata, decreasing in size towards foot. Anus cleioproctic, within right second arch. Gonopore among cerata of anteriormost arch on right.
Anatomy. Masticatory borders smooth ( Fig. 4 View FIGURE 4 E). Radular formula 33 x 0.1.0 ( CASIZ 181357, 5 mm). Radular teeth slightly bilobed with 27–36 elongate, acutely pointed denticles on either side of the inconspicuous central notch ( Fig. 4 View FIGURE 4 F). Teeth progressively smaller in posterior region of radula. Oral glands small, fragile, spongy. Oral glands dorso-laterally to buccal bulb. Salivary glands long.
Reproductive system diaulic ( Fig. 6 View FIGURE 6 C). Preampullary duct widening into moderately short ampulla. Postampullary duct dividing into oviduct and vas deferens. Vas deferens entering into wider proximal portion of penial sac with unarmed penial papilla. Receptaculum seminis huge, rounded, short stalk connecting to long oviduct, before latter forms female glands. Vagina ventral to penis.
Remarks. When Baba (1933a) described Baeolidia japonica , he did not mention anything about colouration. It was sixteen years later ( Baba 1949) when the first colour figure of B. japonica was published. In that contribution, Baba depicted this species as “ground-colour dark brown, the whole back sprinkled with opaque white spots”. This description clearly does not match with our specimens, which are mainly white. Nevertheless, in the same area where Baba described B. japonica , both colour types (brownish and whitish) cohabit ( Rudman 2002; Nakano 2004; Ono 2004). Since one of our specimens is from the type locality of B. japonica , we conclude that both colour types are conspecific.
Excluding Baeolidia scottjohnsoni sp. nov., the main internal differences between B. japonica and the remaining species of the genus is the shape and the size of the receptaculum seminis. This structure is bigger and more rounded in B. japonica than in the remaining Baeolidia species. Due to this particular receptaculum seminis, it cannot be confounded with the remaining species of the genus. Its distinction from B. scottjohnsoni sp. nov. will be discussed in the remarks section for that species.
Carmona et al. (2013) studied this species from a molecular approach ( Fig. 1 View FIGURE 1 ).





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Baeolidia japonica Baba, 1933
| Carmona, Leila, Pola, Marta, Gosliner, Terrence M. & Cervera, Juan Lucas 2014 |
Baeolidia japonica
| Baba 1933: 282 |
Spurilla japonica
| Baba 1933 |
Berghia japonica
| Baba 1933 |