
Anthocoris miyamotoi Hiura, 1959
|
publication ID |
https://doi.org/ 10.37520/aemnp.2021.022 |
|
publication LSID |
lsid:zoobank.org:pub:32519CC9-3658-469A-926D-6A1EBEE0FC59 |
|
persistent identifier |
https://treatment.plazi.org/id/03D687AF-A122-FFD3-6B4D-FBFBFC6980E4 |
|
treatment provided by |
Plazi |
|
scientific name |
Anthocoris miyamotoi Hiura, 1959 |
| status |
|
Anthocoris miyamotoi Hiura, 1959
( Figs 8A–D View Fig , 9F View Fig , 11A–B View Fig , 12F View Fig , 14A–C View Fig , 15F View Fig , 17F–G View Fig , 22 View Fig )
Anthocoris miyamotoi Hiura, 1959:3 . Holotype:J, Japan, Tokara Islands , Nakanoshima Is. (OMNH).
Anthocoris miyamotoi : Fඈඋൽ (1979): 56 (listed, distribution); TඈආඈKඎඇං (1981): 109 (record); Mංඒൺආඈඍඈ & YൺඌඎඇൺGൺ (1989): 165 (listed, distribution);TඈආඈKඎඇං (1993):pl. 27, 168 (photo, diagnosis, habitat, distribution);Pඣඋංർൺඋඍ (1996): 113 (catalogue, Palaearctic);Yൺඌඎඇൺ- Gൺ (1997a): 32 (listed); YൺඌඎඇൺGൺ (1999): 22 (listed, distribution); YൺඌඎඇൺGൺ (2001b): 282 (listed); HൺඒൺඌHං (2002): 136 (listed, distribution); TඈආඈKඎඇං & IඌHංKൺඐൺ (2002): 173 (record); TඈආඈKඎඇං & HൺඒൺඌHං (2006): 293 (record);AඎKൾආൺ et al. (2013a): 86 (catalogue); Yൺආൺൽൺ et al. (2016): 422 (catalogue, distribution); YൺඌඎඇൺGൺ et al. (2018): 138 (photo); Uඋൺඒൺආൺ et al. (2019):79, 81–82 (record, photo, habitat); MංඒൺඓൺKං et al. (2020): 65–66 (rearing method).
? Anthocoris miyamotoi (doubtful identifications,not verified): Mංඒൺආඈඍඈ (1965): 96, pl. 48 (diagnosis, habitat, distribution, photo); NൺKൺඃංආൺ (1968): 46 (record); SHංආංඓඎ (1969): 244 (record); Tൺඐൺඋൺ (1970): 67 (record); NൺKൺඃංආൺ (1976): 97 (record); Mංඒൺඍൺ et al. (1977): 566 (record); IඐൺඌൺKං (1983): 14 (record); TൺKൺKඎඋൺ & Tඌඎඍඌඎආං (1983): 178 (record); TൺKൺKඎඋൺ (1984): 6 (record); Sൺඌൺඃං (1985): 58 (record); OඍඌඎKൺ & YඈඌHංඓൺKං (1987): 596 (record); OඍඌඎKൺ & AඋൺආൺKං (1989): 29 (record); IർHංඍൺ (1988): 132 (record); AඌൺඈKൺ & IൾKං (1990): 155 (listed, distribution); Lൾൾ & Kඐඈඇ (1991): 13 (record); IർHංඍൺ & SඎඓඎKං (1993): 199 (distribution); Lൾൾ et al. (1993): 10 (record); Lൾൾ & Kඐඈඇ (1994): 63 (listed); Kඐඈඇ (1994): 177 (listed); Kඐඈඇ et al. (1996a): 109 (listed); Kඐඈඇ et al. (1996b): 453 (record); HൺඒൺඌHං (1998): 165 (record); IർHංඍൺ (1998): 27 (record); KංඌHංආඈඍඈ et al. (1998): 84 (distribution); Hඎൺ (2000): 198 (listed, distribution); IർHංඍൺ et al. (2000): 110 (record); Yൺඇඈ & Yൺආൺආඈඍඈ (2000): 168 (record); Bඎ & ZHൾඇG (2001): 118, 146 (in key, redescription, figures); Iඐൺඍൾ Pඋൾൿൾർඍඎඋൾ (2001): 179 (listed); Kඐඈඇ et al. (2001): 80 (catalogue, record, distribution); YൺඌඎඇൺGൺ & HංGඎർHං (2002):88 (record, associated plant); KൺGൺඐൺ & HංGඎർHං
(2003): 205 (record, distribution); SඎඓඎKං (2003): 147 (distribution);
HൺඒൺඌHං & OඓൺKං (2004): 223 (distribution); HൺඒൺඌHං & IඌHංKൺඐൺ
(2005): 94 (photo of fifth instar larva and adult, diagnosis, habitat,
prey, distribution); WൺർHං (2006): 66 (record); TൺGඈ (2006): 29
(record); Eඇඃඎ (2007): 89 (distribution); Kൾ & Bඎ (2007): 92–93
(figure, description of female genitalia); Mංඒൺආඈඍඈ (2008): 164, pl.
57 (diagnosis, habitat, distribution, photo); IർHංඍൺ (2009): 68 (listed);
AඈKං (2010): 72 (record); OGൺඐൺ et al. (2012): 8 (record); Yൺඇඈ et al. (2012): 87 (record); Hൺඈ & Mൺ (2013): 36 (listed); JඎඇG et al.
(2013): 422 (catalogue, diagnosis, distribution); NඈඓൺKං & NඈඓൺKං
(2013):33 (record); ZHൾඇG & Lංඇ (2013):298 (redescription, photo);
SඎඓඎKං (2014b): 7 (record); MൺൾHൺඋൺ (2015): 29 (record, habitat,
phenology); NඈඓൺKං et al. (2015): 18 (record); Kඈආൺඍඌඎ (2016): 97
(listed); NඈඓൺKං et al.(2016): 82 (record); JඎඇG & Lൾൾ (2017):37–38,
71 (diagnosis, redescription,distribution, photo, figure); HൺඒൺඌHං et al.
(2018): 182 (distribution); HൺඒൺඌHං & KൺൽඈඐൺKං (2019): 19 (listed);
TൺඇൺKൺ (2019):64, 111 (listed); Kඈආൺඍඌඎ & HංൽൺKൺ (2020):108–109
(record, photo); Iඍඈ et al. (2020): 113 (record); OKඎඓඈඇඈ & Yൺආൺආඈ-
ඍඈ (2020): 82 (record); SHංඓඎඈKൺ Pඋൾൿൾർඍඎඋൾ (2020): 128 (listed). Type material examined. Hඈඅඈඍඒඉൾ: J ( Figs 8A–B View Fig ), ‘Osaka Mun.Mus. \ Tokara Is. Exp. \ Nakanoshima \ 3–13. vi. 1953 \ T. Nakane’ [printed], ‘ HOLOTYPE J [handwritten] \ Anthocoris [handwritten] \ miyamotoi [handwritten] \ HIURA, 1959 [handwritten] \ I. HIURA Det. [printed]’, ‘OMNH TI 189’ [handwritten] (OMNH). Additional material examined. JAPAN: HඈඇඌHඎ: Chiba Pref.: 2 JJ 1 ♀, Tateyama-shi, Mera, 15.vi.2002, M. Tomokuni (NSMT). Kanagawa Pref.: 2♀♀, Yokosuka-shi, Sarushima, 7.viii.1999, S.Arai (TKPM); 1♀, Yokosuka-shi, Mt. Take-yama, 12.ii.1990, K. Aoki (TKPM). Mie Pref.: 1 ♀, Shima-shi, Daiô-cho, Tomoyama-Nakiri, 11.viii.1996, T. Konishi (EUM). Shiga Pref.: 8 ♀♀, Ôtsu-shi, Seta-gawa Riv., 28.vii.2004, K. Yamada (TKPM). Hyogo Pref.: 2 ♀♀, Minamiawaji-shi, Nada-Yoshino, 25.v.2017, H. Inoue (TKPM). Wakayama Pref.: 1 J ( Fig. 8C View Fig ), Kushimoto-cho, Hashikui, 22.xi.2001, K.Yamada (TKPM). Yamaguchi Pref.: 2 ♀♀, Hikari-shi, Mt. Senbou-san, 18.vii.2010, T. Ban (TKPM); 2 JJ, Kaminoseki-cho, Nagashima Is., 23.–25.vi.2000, S. Nagashima (TKPM). Iඓඎ Iඌඅൺඇൽඌ: Ôshima Is.: 1♀, 17.iv.1974,Y.Furuki (EUM).Miyake-jima Is.: 2 ♀♀, 15.v.1999, T. Kishimoto (TKPM). Hachijô-jima Is.: 4 JJ 10 ♀♀, Mt. Hachijôfuji, 560–850 m, 2.–5.vii.2001, M.Tomokuni (NSMT); 2 JJ 1 ♀, same locality, 3.viii.2001, T. Ishikawa (TKPM); 1 J, Mt. Mihara-yama, 24.vii.2001, J. Ogawa (EUM). SHංKඈKඎ: Tokushima Pref.: 1 ♀, Tokushima-shi, Ichinomiya-cho, 20.iii.2017, T. Nakanishi (TKPM); 1 J 2 ♀♀, Ôbara-cho, 24.v.2016, T. Nakanishi (TKPM); Sanagouchi-son, Ozakai, 34.005000N 134.478889E: 1 ♀, 16.x.2013; 1 ♀, 27.xii.2013; 1 J, 18.v.2015; 2 JJ 4 ♀♀, 7.x.2016, T. Nakanishi (all in TKPM); 5 JJ 2 ♀♀, Kamiyama-cho, Orono, 33.987778N 134.398611E, 13.v.2016, T. Nakanishi (TKPM); Katsuura-cho, Nue, 33.940278N 134.541944E: 1 J, 29.iv.2013; 2 JJ, 14.v.2013; 3 JJ, 30.v.2013; 1 ♀, 25.ii.2014; 3 JJ 3 ♀♀ (one in Fig. 15F View Fig ), 26.iv.2016; 1 J 2 ♀♀, 8.v.2016; 1 J 2 ♀♀, 21.v.2017, T. Nakanishi (all in TKPM); 1 ♀, Katsuura-cho, Hoshidani, 29.x.2013, T. Nakanishi (TKPM); 3 JJ 7 ♀♀ (one in Fig. 9F View Fig ), Anan- -shi, Kuwano-cho, 33.870833N 134.610278E, 29.iv.2016, T. Nakanishi (TKPM); 1♀, Kaiyô-cho,Kôno, 6.vi.2012, K.Yamada (TKPM). Kagawa Pref.: 1 ♀, Shôdoshima Is., Tonosyô-chô, Oobe, 27.iii.2020, G. Kisaki (TKPM). Ehime Pref.: 1J 1♀, Matsuyama-shi, Kashima, 29.iii.1953, M. Miyatake (EUM); 1 J, Matsuyama-shi, Takahama-machi, 14.v.2002, T. Kurihara (EUM). Kochi Pref.: 3♀♀, Tôyô-cho,None, 3.v.2000, M.Takai (TKPM); 1♀, Tosashimizu-shi,Ashizuri Cape, 25.–26.vii.1967,M.Miyatake et al. (EUM); 1 ♀, same locality, 23.v.1983, M. Miyatake (EUM); 1 ♀, Tosashimizu-shi, Ashizuri-hantô, 29.vi.2001, M. Takai (TKPM). KඒඎඌHඎ: Fukuoka Pref.: 1 ♀, Sasaguri-machi, Mt. Wakasugi-yama, 1.iv.1959, Y.Miyatake (OMNH); 1♀, Fukuoka-shi, Hakozaki, 17.iv.1958, Y. Miyatake (OMNH); 1 ♀, Fukuoka-shi, Hirao, 29.v.1959, Y. Miyatake (OMNH). Nagasaki Pref.: 1 ♀, Nagasaki-shi, Kônoura, 4.viii.1996, T. Yasunaga (TKPM); 1 ♀, Nagasaki-shi, Nomozaki, 7.iv.1978, M. Sakai (TKPM); 1 ♀, Nagasaki-shi, Nomozaki, Kabashima Is., 32.5555N 129.7768E, male flower of Mallotus japonicus , 24.viii.2013, T.Yasunaga (TYCN); 1 J, Nagasaki-shi, Kawaguchi-Park, 32.76685N 129.86355E, under flakes of bark of Zelkova serrata , 2.ii.2018, T. Yasunaga et al. (NWHS); 1 ♀, Nagasaki-shi, 4.iv.1973, Y. Furuki (EUM); 1 J ( Figs 11A–B View Fig ), Nagasaki-shi, 24.iv.2012, T. Yasunaga (TKPM); 1 ♀, Saikai- -shi, Seihi-cho, Nagasaki Biopark, 32.988073N 129.78233E, Ligustrum japonicum inflorescens, 29.vi.2013, T. Yasunaga (TYCN). Kagoshima Pref.: 2 JJ (one in Fig. 8D View Fig ) 5 ♀♀, Satsumasendai-shi, Kashima- -cho, Imuta, 24.iv.2012, H. Inoue (TKPM); 1 ♀, Minamiôsumi-cho, Ôdomari, 19.iii.1973, M. Miyatake (EUM). TඌඎඌHංආൺ Iඌඅൺඇൽ: 1 ♀, Kamiagata-cho, Saozaki Park, 15.–16.vii.2000, J. Ogawa (EUM); 1 J, Kamiagata-cho, Mt. Mitake, 15.vii.2000, J. Ogawa (EUM); 1 ♀, Toyotama-cho, Waita, 30.vi.2008, T. Nakayama (TKPM). RඒඎKඒඎ Iඌඅൺඇൽඌ: Yakushima Is.: 1 J, Miyanoura, 16.iv.1954, Y. Kurosawa (TKPM); 2 JJ 2 ♀♀, Onoaida, 3.i.1965, I. Hiura (OMNH). Kuchinoshima Is.: 1 J, 21.v.1962, M. Sato (EUM). Nakanoshima Is.: 4 JJ (one in Figs 12F View Fig , 14A–C View Fig ) 1♀ 1 unsexed, Takamoto-rindô, 6.–10.vi.2005,T. Mita (TKPM); 1 J, 2.v.1971, M. Sakai (EUM). Amami-Ôshima Is.: 1 ♀, Tatsugô-cho, 24.iii.2011, S. Kudo (TKPM); 2 JJ 4 ♀♀, Tatsugô-cho, 17.iv.2016, S. Kudo (TKPM); 1 ♀, Amami-shi, Daikuma-cho, 10.iv.1971, M. Sakai (EUM); 1♀, Amami-shi, Mt.Ogami-yama, 30.iv.1977,A. Oda (NSMT); 7 JJ 4 ♀♀, Amami-shi, Asado, 31.v.1993, T. Yasunaga (TKPM); 1 ♀, Chûou-rindô, 18.vi.1995, Y. Sawada (TKPM); 1♀, Amami-shi, Sumiyô, Hatsuno-bashi, 5.vii.2005, light trap, T. Ishikawa (TKPM); 1 J 2 ♀♀, Yamato-son, Mt. Yuwan-dake, 17.–19.iv.1971, M. Sakai (EUM); 1 ♀, same locality, 30.v.1993, T. Yasunaga (TKPM); 2 ♀♀ 1 unsexed, Setouchi-cho, Mt. Yui-dake, 24.iv.1997, T. Ishikawa (TKPM); 1 J, 25.v.2004, T. Mita (TKPM); 1 ♀, Ikari, 6.v.1960, T. Shibata (OMNH). Okinawa-hontô Is.: 1 ♀, Kunigami-son, Mt. Nishime-dake, 25.v.1993, T. Yasunaga (TKPM); 1 J, Kunigami-son, Sate, 24.v.1993, T. Yasunaga (TKPM); 2 ♀♀, Kunigami-son, Yona, 20.v.1993, T. Yasunaga (TKPM); 1 ♀, 12.iv.1996, Y. Nakatani (TKPM); 2 JJ 1 ♀, 18.v.2002, K. Yamada (TKPM); 1♀, Kunigami-son, Mt.Terukubi-yama, 23.v.1993, T.Yasunaga (TKPM); 2 ♀♀, Kunigami-son, Hentona, Shinrin Park, 31.iii.2001, K. Yamada (TKPM); 1 ♀, Kunigami-son, Mt. Yonaha-dake, 13.vi.2003, T. Tsuru (TKPM).
Differential diagnosis. Recognized by the following combination of characters: head and callus of pronotum orangish brown to reddish brown ( Figs 8A–B View Fig ); clavus with basal and apical portions blackish brown, inner margin narrowly darkened; endocorium with basal and apical portions mostly blackish brown; embolium with basal and apical portions blackish brown; median portion of clavus and outer part of median portion of endocorium and remaining area of embolium whitish and subhyaline; area in front of cuneal fracture narrowly whitish or subhyaline; outer (posterior) margin of ostiolar peritreme touching or close to supracoxal area ( Fig. 9F View Fig ); paramere ( Figs 11A–B View Fig , 14B–C View Fig ) sickle-shaped, widened at basal 2/3, apical 1/3 needle-like, with distinct longitudinal groove; and copulatory tube ( Fig. 15F View Fig ) approximately 0.25 mm in length, basal 2/3 very thick and wrinkled and then rapidly becoming narrow apically. Most closely resembling A. venustus sp. nov. in general appearance but distinguished from that species by the orangish brown to reddish-brown head and pronotal callus, sickle-shaped and rather widened paramere, and a shorter copulatory tube, at the basal 2/3 of which is very thickened and wrinkled. It is also similar to A. dividens Bu & Zheng, 2001 from China, but clearly distinguished by the yellowish-brown antennal segment II, which has the base and apical 1/4 to 1/3 dark brown (in A. dividens , uniformly blackish brown), membrane with basal and innermost portions and area behind apex of cuneus off- white (in A. dividens , membrane with grayish-white V-shaped marking on basal part), and sickle-shaped paramere (in A. dividens , lamellate).
Redescription. Coloration. Head ( Figs 8A–D View Fig ) reddish brown to blackish brown; eyes reddish black; margin of ocellus red to reddish brown. Antennal segment I dark brown, sometimes yellowish brown on basal half; segment II yellowish brown, dark brown on base and apical 1/4 to 1/3; segment III yellowish brown, dark brown on apical half; segment IV dark brown. Labium ( Fig. 8B View Fig ) uniformly black to blackish brown. Pronotum ( Figs 8A–D View Fig ) generally blackish brown; callus usually orangish brown to reddish brown, sometimes mostly blackish brown but reddish tinge at median part. Scutellum overall black to blackish brown. Clavus with basal and apical portions blackish brown, inner margin narrowly darkened ( Figs 8A,C–D View Fig ); endocorium with basal and apical portions mostly blackish brown ( Figs 8A,C–D View Fig ); embolium with basal and apical portions blackish brown ( Figs 8A,C–D View Fig ); cuneus wholly blackish brown; median portion of clavus and outer part of median portion of endocorium and remaining area of embolium whitish or subhyaline ( Figs 8A,C–D View Fig ); area in front of cuneal fracture narrowly whitish or subhyaline; membrane smoky dark brown, with basal and innermost portions and area behind apex of cuneus off- white ( Figs 8A,C–D View Fig ). Legs yellowish brown to reddish brown; trochanters tinged with yellowish brown; apical half or apical 2/3 of tibiae sometimes paler. Venter of the thorax reddish brown to blackish brown. Abdomen black to blackish brown. Pygophore blackish brown with light brown paramere.
Structure. Body ( Figs 8A–D View Fig , 17G View Fig ) elongate, shiny on dorsal and ventral surfaces, sparsely covered with suberect, simple, yellowish setae. Head ( Figs 8A–D View Fig ) cylindrical, impunctate, approximately as long as width across eyes; three pairs of long, erect trichobothria on dorsal surface of head, one on anterior clypeus, one in front of eye, and one behind ocellus; anteocular region slightly longer than length of eye in dorsal view; vertex approximately twice as wide as eye in dorsal view in male, approximately 2.2 times as wide as eye in female; postocular region constricted, demarcated by transverse shallow furrow; neck long, smooth, highly polished; eye oblong, not exceeding level of dorsal surface of head in lateral view in both sexes, but exceeding level of ventral surface of head in male and not exceeding in female. Antennae ( Figs 8A–D View Fig ) densely covered with short, decumbent, yellowish setae interspersed with long, erect setae, of which the longest are as long as or slightly shorter than width of corresponding segment; segment I barely reaching apex of head, sparsely covered with short setae; segment II gradually thickened toward apex, a little thicker in the male than in the female, approximately as long as head width across eyes; segment III approximately 0.6–0.7 times as long as segment II; segment IV longer than segment III. Labium ( Fig. 8B View Fig ) just reaching procoxae; segment III approximately 4.0 times as long as segment II; segment IV approximately 0.58 times as long as segment III.
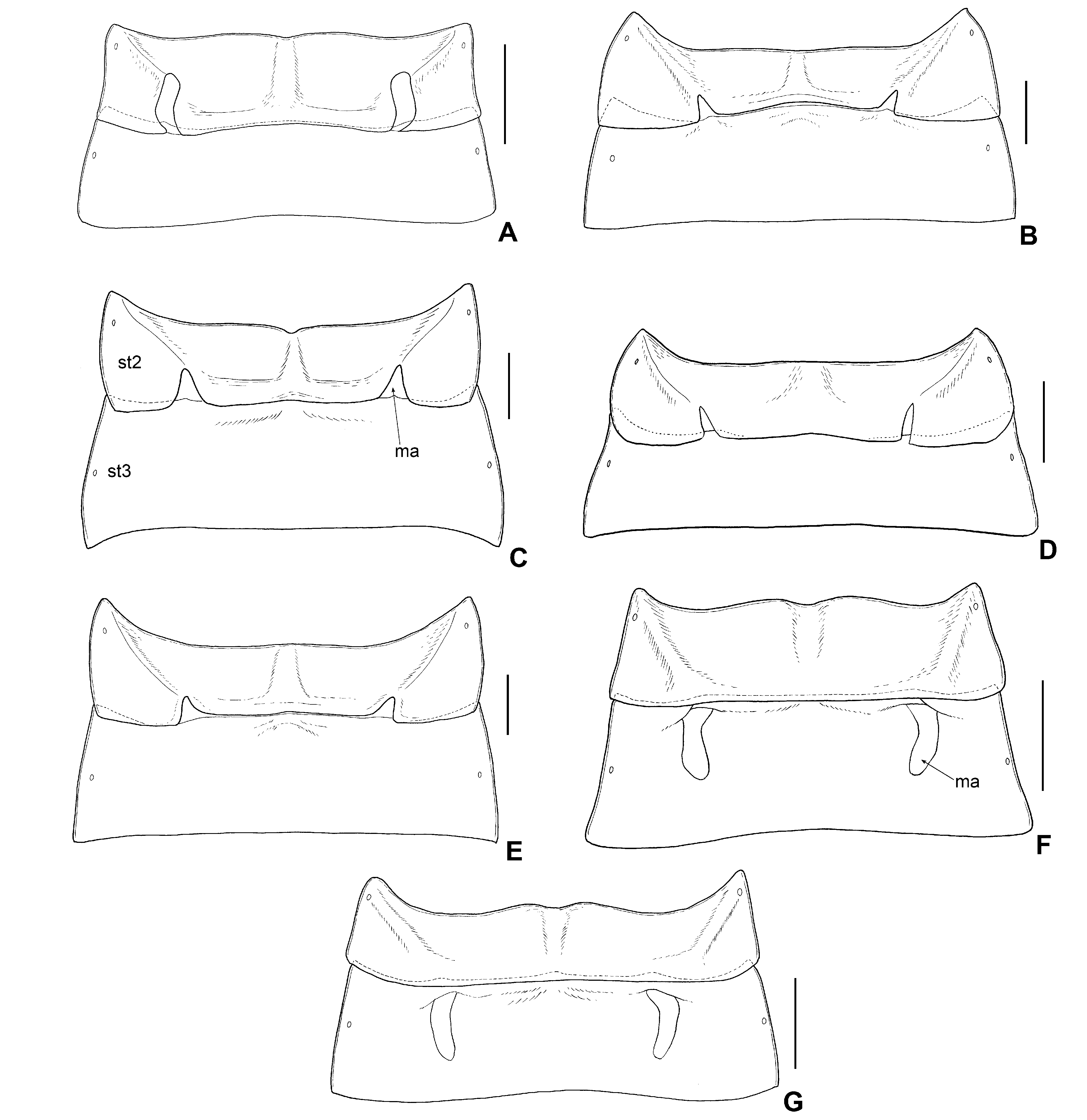
Pronotum ( Figs 8A,C–D View Fig ) with pairs of long, erect trichobothria on midline of collar and on each anterior and posterior angle; anterior margin very slightly curved, approximately 0.7 times as long as mesal length; lateral margin slightly sinuate; lateral carina slightly expanded anteriorly, more obscure posteriad; posterior margin emarginate, approximately 2.5 times as wide as anterior margin; collar long, transversely rugose, with scattered long setae; callus extremely swollen, polished, impunctate, sides strongly bulging, demarcated posteriorly by deep impression; deep impression with short parallel carinae and coarse punctures; posterior lobe behind callus densely covered with minute punctures. Scutellum ( Figs 8A,C–D View Fig ) smooth, sub-equilateral, wider at base than long, depressed through middle. Hemelytra ( Figs 8A–D View Fig ) sparsely covered with long, erect, simple, yellowish setae, and with minute punctures; costal margin slightly sinuate; maximum width of endocorium approximately 1.6 times that of maximum width of embolium; cuneal margin approximately 0.6 times as long as embolial margin; membrane with three veins, outermost vein distinct and slightly curved, inner two veins obscure and very slightly curved. Ostiolar peritreme ( Fig. 9F View Fig ) wide, gently curved anteriorly, narrowing toward apex, and then continuing to a fine carina that reaches the anterior margin of metapleuron; outer (posterior) margin of ostiolar peritreme touching or close to supracoxal area; evaporatorium sparsely covered with long, recumbent setae. Fossula spongiosa present on apex of all tibiae, enlarged on protibiae and small on meso- and metatibiae; apical 1/4 of metatibiae bearing small spines on ventral side.Abdominal sternum III ( Fig. 12F View Fig ) with a pair of banana-shaped membranous areas on anterior half, membranous area slightly constricted at middle; dorsal laterotergites not fused with mediotergites on abdominal segments II and III.
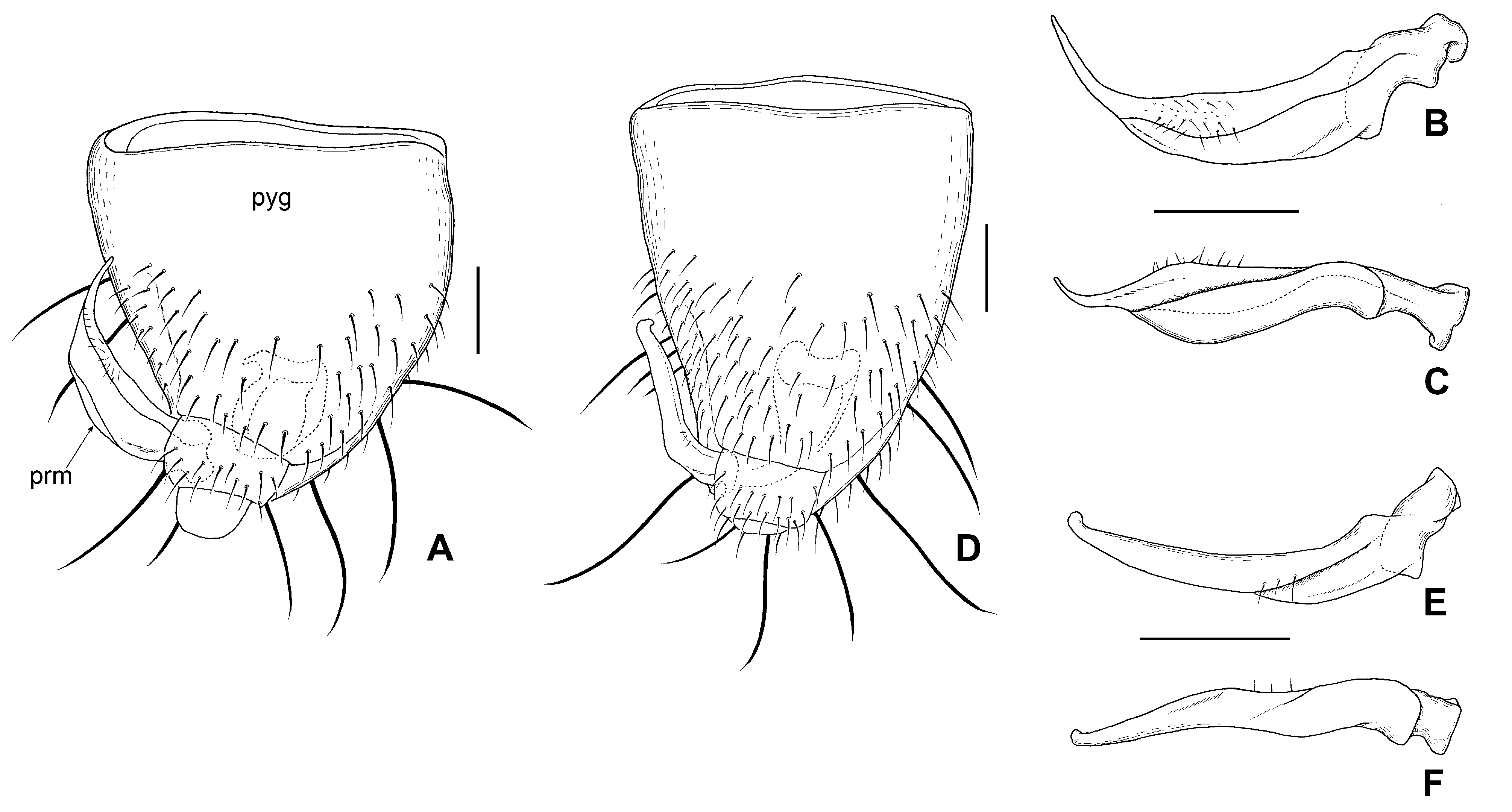
Male genitalia ( Figs 11A–B View Fig , 14A–C View Fig ): Pygophore ( Fig. 14A View Fig ) rather pointed apically, asymmetrical, covered with 6–8 long, stout setae intermixed with short, suberect setae along outer margin and on posteroventral surface, of which the longest are approximately half length of pygophore; mid-dorsal surface hirsute with suberect setae; paramere ( Figs 11B View Fig , 14B–C View Fig ) sickle-shaped, widened at basal 2/3, needle-like at apical 1/3, with several setae on subapical portion, apex exceeding half of left outer margin of pygophore; longitudinal groove distinct, formed by inflection of anterior and posterior edge of paramere.
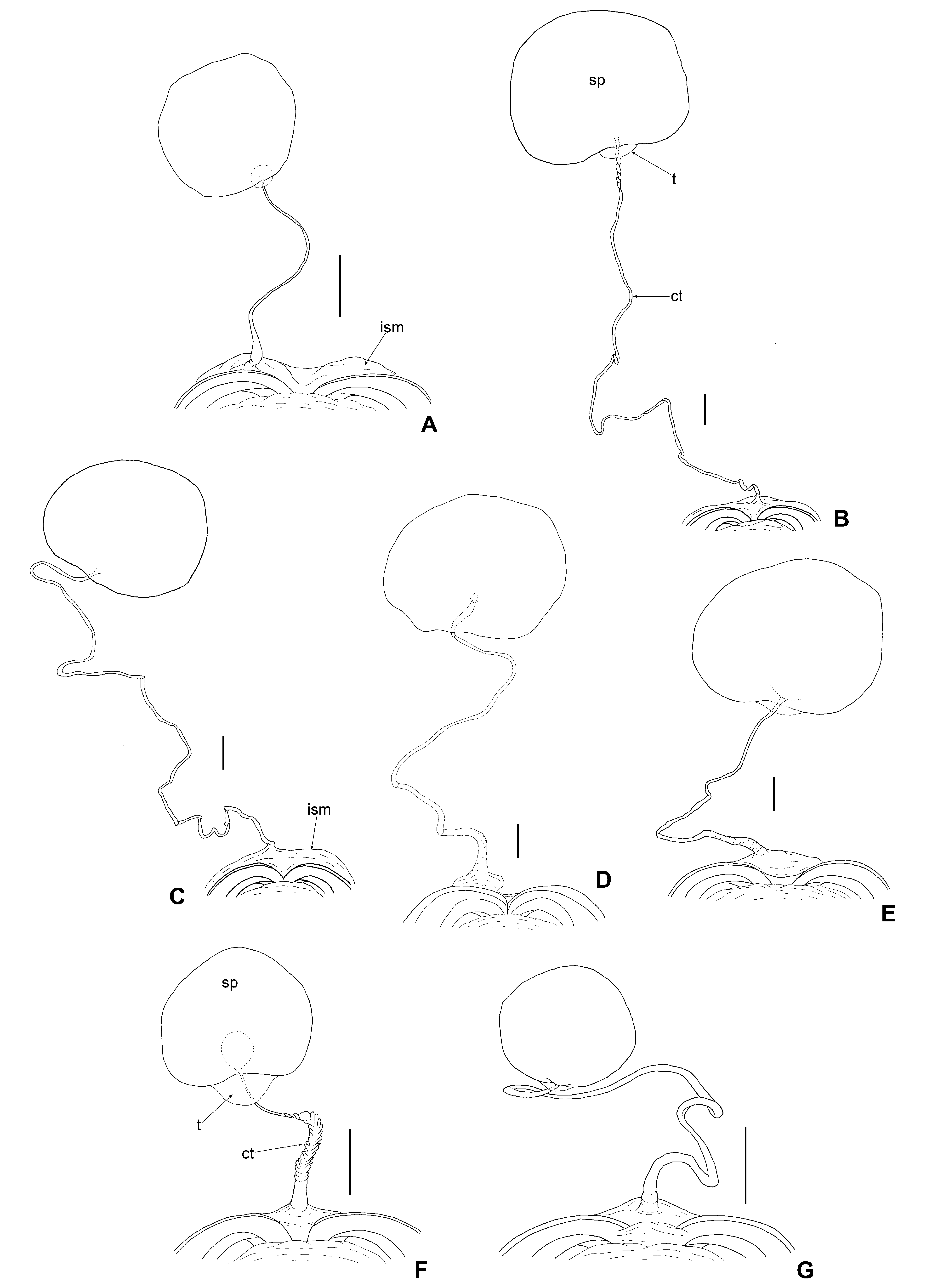
Female genitalia ( Fig. 15F View Fig ): Copulatory tube fused on middle of intersegmental membrane between sterna VII and VIII, approximately 0.25 mm in length, very thick and wrinkled at basal 2/3 and then rapidly becoming narrow apically; trunk of conductive tissue strongly pronounced.
Measurements [mm; JJ (n = 10) / ♀♀ (n = 10)]. Body length 3.08–4.00 / 3.15–4.20; head length (excl. neck) 0.44–0.49 / 0.44–0.52; head width across eyes 0.47–0.55 / 0.48–0.56; vertex width 0.24–0.29 / 0.27–0.30; length of antennal segments I – 0.13–0.17 / 0.14–0.19, II – 0.46–0.59 / 0.43–0.55, III – 0.30–0.37 / 0.29–0.36, and IV – 0.37–0.42 / 0.36–0.41; length of labial segments II – 0.12–0.15 / 0.10–0.17, III – 0.50–0.56 / 0.50–0.62, and IV – 0.28–0.33 / 0.30–0.35; anterior pronotal width 0.38–0.44 / 0.39–0.45; mesal pronotal length 0.52–0.60 / 0.52–0.62; basal pronotal width 0.89–1.10 / 0.94–1.18; length of embolial margin 0.90–1.20 / 0.90–1.25; length of cuneal margin 0.56–0.75 / 0.57–0.79; maximum width across hemelytra 0.94–1.16 / 0.97–1.27.
Bionomics. No specific plant associations are known for Anthocoris miyamotoi , as it is collected from various deciduous and evergreen broadleaf trees. The species is found on Salix spp. , Quercus spp. , Castanopsis spp. , Zelkova serrata , Mallotus japonicus (L. f.) Müll. Arg., Eriobotrya japonica (Thunb.) Lindl. , Elaeagnus macrophylla Thunb. , Rhus javanica L. var. chinensis (Mill.) T. Yamaz. , Schima wallichii (DC.) Korth. subsp. noronhae (Reinw. ex Blume) Bloemb. , and Albizia julibrissin Durazz. Additionally , this species was collected from different parts of the same tree, such as flowers, leaves, stems, peduncles, and bud scales; therefore, it may walk around very actively and does not seem to stay on one particular part of a tree.
In Tokushima, eastern Shikoku, this species is frequently found on Eriobotrya japonica (Rosaceae) , where it preys on the psyllid Cacopsylla biwa Inoue, 2014 ( Hemiptera : Psyllidae ), a serious pest of the loquat. Both nymphs and adults of A. miyamotoi are usually observed on the stems, peduncles, and bud scales, and are present for two months, from early April to the end of May. Anthocoris miyamotoi is easily found on loquat when C. biwa is present, but often disappears from the tree when the psyllid is not abundant. From the end of May to June, A. miyamotoi regularly disappears from loquat, as a consequence of the seasonal decrease in psyllid numbers. Presumably, A. miyamotoi moves between various broad-leaved trees for breeding or when seeking food. Individuals are observed again on loquat in February. Thus, after departing loquat in summer, individuals of the next generations apparently return in autumn or winter, and overwinter there.
In Nagasaki, south-western Kyushu (belonging to warm-temperate climatic zones), this anthocorid seems to complete two or three generations per year, and the adults have been found throughout the year. During winter (December‒February), adults and late instar immature forms were observed to co-occur and hibernate under bark flakes of Zelkova serrata , sometimes together with Anthocoris japonicus (Uඋൺඒൺආൺ et al. 2019) ( Fig. 17F View Fig ). The adults appear to be active even in winter when temperature is high ( Fig. 17G View Fig ).
Based on the above, no specificity for associated plant or specific overwintering sites were recognized in A. miyamotoi . Although the prey preference of A. miyamotoi has been studied insufficiently, this species is certainly a polyphagous predator, and its frequency of occurrence seems to be affected by the amount of food resources.
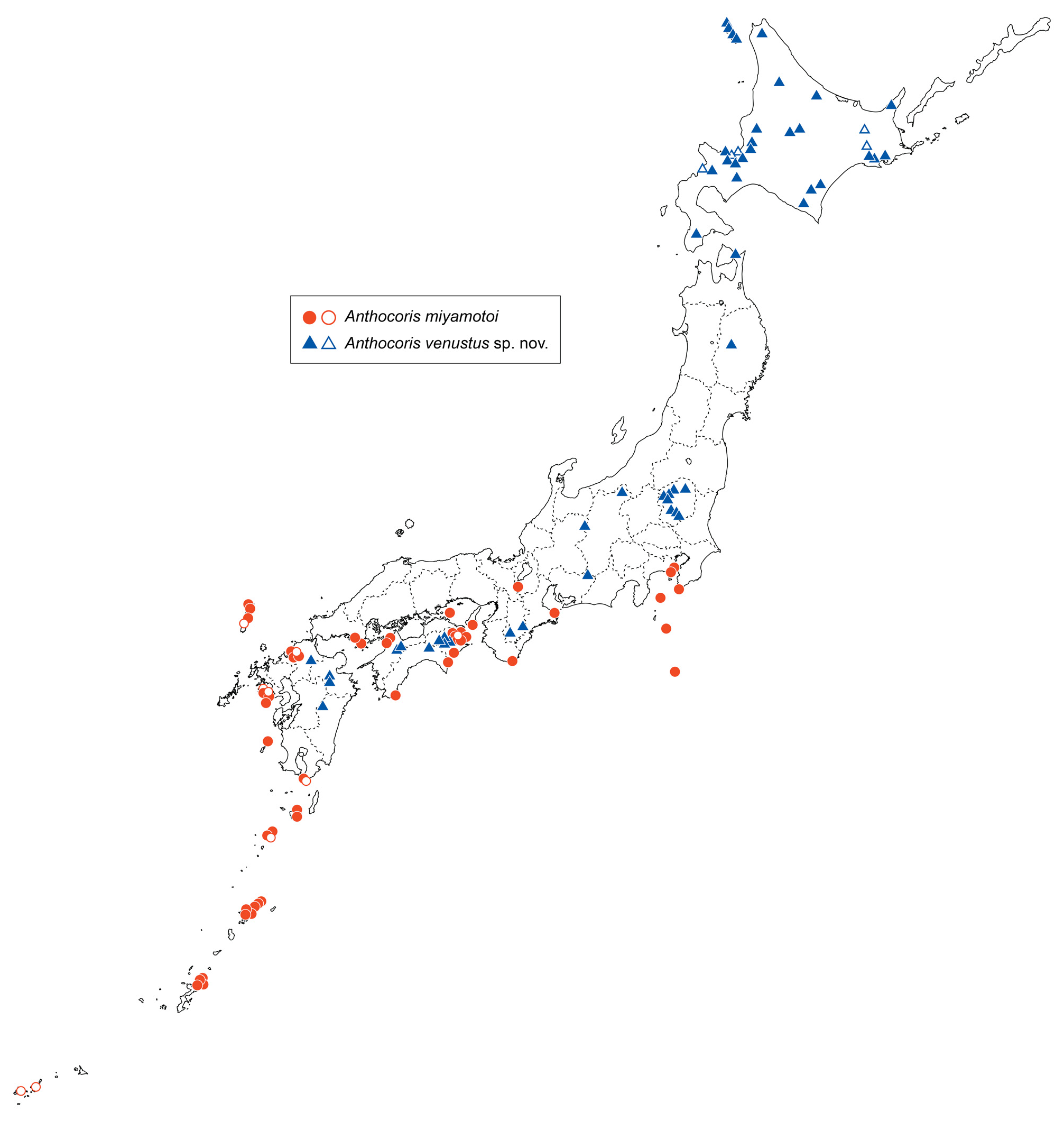
Distribution. Japan: Honshu: Chiba (TඈආඈKඎඇං & HൺඒൺඌHං 2006, material verified), Kanagawa; Shizuoka (TඈආඈKඎඇං 1981, material verified), Mie, Shiga, Wakayama, Hyogo, Yamaguchi; Izu Islands: Ôshima Is., Miyake-jima Is., Hachijô-jima Is. (TඈආඈKඎඇං & IඌHංKൺඐൺ 2002, material verified); Shikoku: Tokushima (Hංඎඋൺ 1959, material verified), Kagawa, Ehime, Kochi; Kyushu: Fukuoka (Hංඎඋൺ 1959, material verified), Nagasaki (YൺඌඎඇൺGൺ et al. 1989, YൺඌඎඇൺGൺ 1997a, material verified), Kagoshima (Hංඎඋൺ 1959, material verified); Tsushima Island (YൺඌඎඇൺGൺ et al. 1989); Ryukyu Islands: Yakushima Is., Kuchinoshima Is., Nakanoshima Is. (Hංඎඋൺ 1959, material verified), Amami-Ôshima Is. (TඈආඈKඎඇං 1993), Okinawa-hontô Is. (HൺඒൺඌHං 2002), Ishigaki-jima Is. (HൺඒൺඌHං 2002), Iriomote-jima Is (HൺඒൺඌHං 2002) ( Fig. 22 View Fig ). The specimens, including previously published ones from Ishigaki-jima and Iriomote-jima Islands, were not examined in the present study; however, the species is distributed on both islands. Based on the specimens verified, this species is currently distributed along the coastal areas of Honshu west of the Kanto district, Shikoku, Kyushu, and the Ryukyu Islands, as well as some small islands (Izu Islands and Tsushima Island) ( Fig. 22 View Fig ).
Remarks. Anthocoris miyamotoi is known to occur in Japan, Chishima (Kuril) Islands (Shikotan Is., Kunashir Is., Iturup Is.), Russia (Primorsky Kray, Sakhalin), Korea (Gyeonggi-do, Gyeongsangbuk-do, Gyeongsangnam-do, Jeju-do), China (Shaanxi), and Taiwan (e.g., KൾඋඓHඇൾඋ 1988, Pඣඋංർൺඋඍ 1996, Bඎ & ZHൾඇG 2001, KൺඇඒඎKඈඏൺ & MൺඋඎඌංK 2006, ZHൾඇG & Lංඇ 2013, JඎඇG & Lൾൾ 2017). During this study, however, our careful examinations based on the holotype of A. miyamotoi revealed that the previous records of this species, at least from Japan, included some misidentifications with A. venustus sp. nov. The records from the Chishima Islands and Russian Far East should probably be assigned to A. venustus sp. nov. based on their distribution. Judging from the figures of Bඎ & ZHൾඇG (2001: p. 145, fig. 220) and JඎඇG & Lൾൾ (2017: p. 71, fig. 98), the shape of the paramere somewhat resembles that of A. venustus sp. nov.; therefore, A. miyamotoi as studied by Bඎ & ZHൾඇG (2001) and JඎඇG & Lൾൾ (2017) is presumed to represent A. venustus sp. nov., or an undescribed species. Chinese and Korean records of A. miyamotoi require careful verification based on their genital structures. The Taiwanese record is probably a misidentification because the habitus photo of ZHൾඇG & Lංඇ (2013: p. 298) is different from specimens collected in Japan in the coloration of hemelytra. This species appears to be an undescribed species closely related to A. miyamotoi .
Hංඎඋൺ (1959) noted that ‘Specimens from Tokara Is. and lower land of Kyushu are light, from mountainous region of Shikoku and Hokkaido are blackish, some of from Kyushu are of intermediate colour pattern’. Examination of the type series of A. miyamotoi specimens from Tokara Islands and the lowland of Kyushu has proven them to be A. miyamotoi , while the blackish specimens of the mountainous region of Shikoku and Hokkaido are shown to be A. venustus sp. nov. Among Hiura’s type series, specimens from Mt. Hikosan, Fukuoka Prefecture were assigned to A. venustus sp. nov., but the remainder of the paratypes from Kyushu were all A. miyamotoi . Thus, the intermediate color pattern of this species as indicated by Hංඎඋൺ (1959), includes A. venustus sp. nov. Remarkable color variation, particularly in the head and pronotum, is seen in both species ( Figs 8 View Fig , 16F View Fig , 17G View Fig ); it is therefore difficult to identify the species accurately by coloration alone, and the genital structures are currently the only reliable character for clear determination of the species ( Fig. 14 View Fig ).





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Anthocoris miyamotoi Hiura, 1959
| Yamada, Kazutaka & Yasunaga, Tomohide 2021 |
Anthocoris miyamotoi
| HIURA I. 1959: 3 |