
Paludocyon, Morales & Fejfar & Heizmann & Wagner & Valenciano & Abella, 2021
|
publication ID |
https://doi.org/ 10.37520/fi.2021.011 |
|
persistent identifier |
https://treatment.plazi.org/id/0385BC16-FFFE-FFDE-FC34-FE9DFB154C05 |
|
treatment provided by |
Felipe |
|
scientific name |
Paludocyon |
| status |
gen. nov. |
Paludocyon bohemicus ( SCHLOSSER, 1899)
Text-figs 1–4 View Text-fig View Text-fig View Text-fig View Text-fig , Tabs 1a, 2a
1861 Amphicyon intermedius Mey.; Suess, p. 224, pl. II.
1868 Amphicyon intermedius H. v. Meyer; Peters, p. 190.
1899 Pseudocyon bohemicus n. sp.; Schlosser, p. 124, pl. XIV, figs 3, 4, 9.
1901 Amphicyon bohemicus Schlosser ; Schlosser, p. 62, pl. I, figs 18, 23–26, 29, 30.
1929 Amphicyon bohemicus Schloss. ; Viret, p. 113.
1965 Amphicyon steinheimensis bohemicus (Schlosser) 1899 ; Kuss, pp. 40–46, figs 16–23.
1973 Amphicyon bohemicus ; Heizmann, pp. 17–20.
1977 “ Amphicyon ” bohemicus ; Ginsburg, pp. 94–95.
1999 Cynelos (Heizmannocyon) bohemicus ( SCHLOSSER, 1899) ; Ginsburg, p. 116.
2000a Cynelos (Heizmannocyon) bohemicus (Schlosser) 1899 ; Ginsburg, p. 11.
2003 Amphicyon bohemicus ; Hunt, p. 105.
2008 C [ynelos] bohemicus ; Peigné et al., p. 954.
2015 Cynelos bohemicus ; Hunt and Stepleton, p. 2.
2016 Amphicyon bohemicus ( Schlosser 1899) ; Fejfar and Heizmann, p. 320, figs 9, 10, 11.1–5+11–17.
2020 Cynelos cf. bohemicus ( Schlosser 1899) ; Jiangzuo et al., p. 23, fig. 4.4–6.
L e c t o t y p e. NM-Pv 11677, left m1, figured by Schlosser (1899: pl. 14, fig. 4; Text-fig. 4a View Text-fig herein). Lectotype designated by Kuss (1965: 41; he listed it under the old number 4330).
P a r a l e c t o t y p e s. NM-Pv 11678 (old no. 4329), left P4, figured by Schlosser (1899: pl. 14, fig. 3); NM-Pv 11679 right incomplete M2, and NM-Pv 11731 right complete M3 (both old no. 205/1), figured by Schlosser (1899: pl. 14, fig. 3); and NM-Pv 11680 (old no. 4310) left m3, figured by Schlosser (1899: pl. 14, figs 4 and 9).
T y p e l o c a l i t y. Tuchořice, the Czech Republic.
A g e. Early Miocene, MN 3.
D i a g n o s i s. Same as genus.
Additional material from type locality. NM-Pv 11681 ( TU 7391149 ), left P4; NM-Pv 11682 ( TU 7391150 ), left P4; NM-Pv 11683 ( TU 739113), right P4; NM-Pv 11684 ( TU 739155), left P4; NM-Pv 11685 ( TU 739176), right M1; NM-Pv 11686 ( TU 7391150 ), left M1; NM-Pv 11687 ( TU 739185), M2 right; NM-Pv 11688 ( TU 739149), left M2; NM-Pv 11689 ( TU 739122), right M2; NM-Pv 11690 ( TU 7391150 ), left M2; NM-Pv 11691 ( TU 739165), right M3; NM-Pv 11692 ( TU 739185), left M3; NM-Pv 11695 (TU-739157), right mandible with p2–m2; NM-Pv 11697 ( TU 739135), right mandible with p3–m3; NM-Pv 11698 ( TU 739142), association of right p4, m1 and m2; NM-Pv 11699 ( TU 739141), left mandible with p3–m3; NM-Pv 11700 ( TU 739177), left mandible with canine, p1–p2 alveolus and p3–m2; MN-Pv 11722, association of left p4–m3 .
D e s c r i p t i o n. NM-Pv 11678, left P4 (paralectotype, Text-fig. 3a View Text-fig ). Well-developed basal cingulum, especially in the lingual wall. The protocone is located in a mesial position; it is wide but not very prominent or clearly differentiated from the cingulum. The union with the base of the paracone is not rectilinear, marking a clearly pronounced inflection. There is no isolated parastyle, although the mesial crista of the paracone bifurcates before reaching the base of the tooth. One of the branches is directed towards the protocone and the other mesiobuccally. The paracone is tall and pyramidal and the metastyle is relatively long. Both paracone and metastyle display vertical wear facets on the lingual side.
NM-Pv 11681, left P4 ( Text-fig. 3b View Text-fig ). Differs from the former P 4 in which the paracone mesial crista is more pronounced before reaching the basal cingulum. The protocone is somewhat smaller and the mesial inflection between the base of the protocone and the paracone is less marked. NM-Pv 11682, right P4, is similar in morphology to the previous specimen, with a more reduced protocone and without the inflexion at the junction between the bases of the paracone and the protocone. NM-Pv 11683, right P4, has the same morphology to that of the paralectotype (NMPv 11678). NM-Pv 11684, left P4 ( Text-fig. 3c View Text-fig ), is somewhat larger than the other P4s and possesses a particularly wide protocone.
NM-Pv 11685, right M1 ( Text-fig. 3d View Text-fig ). Molar with subtriangular occlusal shape; it is relatively short, with a distally projected narrowed lingual area, moderately developed and with a mesiolingually strong basal cingulum. The paracone is high compared to the metacone. Small parastyle and metastyle. Paraconule and metaconule differentiated from the protocone cristae, almost symmetrical in comparison to the transversal axis of the molar; metaconule larger than paraconule. Very large but low dune-form protocone. Trigone valley sub-rounded.
NM-Pv 11686, left M1 ( Text-fig. 3e View Text-fig ). Differing from the previous M 1 in the greater development of the lingual cingulum, which completely surrounds the protocone, from the base of the paraconule to the base of the metaconule, and also a wider lingual area.
NM-Pv 11687, M2 right ( Text-fig. 3f View Text-fig ). Sub-quadrangular in shape, with a short buccal wall and a wide transverse diameter in comparison with the M1. Moderately high buccal cusp s. Strong buccal cingulum, especially at the base of the paracone. Low protocone forming a single semicircular crista which reaches the base of both the paracone and the metacone. Very strong lingual cingulum, which is semicircular and completely surrounds the protocone. Other M2s such as Pv 11688 ( Text-fig. 3g View Text-fig ), NM-Pv 11689 ( Text-fig. 3h View Text-fig ) and NM-Pv 11690 ( Text-fig. 3i View Text-fig ) are similar in morphology to the previously described M2.
NM-Pv 11691, right M3 ( Text-fig. 3j View Text-fig ), small molar with reduced metacone. Central protocone opposite to the paracone, both are joined by a circular serrated crista. Strong buccal and lingual cingula. The left M3 NM-Pv 11692 ( Text-fig. 3k View Text-fig ) is somewhat smaller than the previous specimen, differing in the complete loss of the metacone.
NM-Pv 11677, left m1 (lectotype, Text-fig. 4a View Text-fig ). Very robust molar, with a short paraconid presenting a vertical mesial cristid. High and very bulky protoconid. Metaconid quite strong, slightly displaced distally and with the lingual wall somewhat rounded. Short talonid, almost completely occupied by the hypoconid, the buccal wall of which is almost vertical. Low and crestiform entoconid. Small hypoconulid. Lingually weak basal cingulum, much stronger buccally, particularly at the base of the hypoconid, which is thickened apically.
NM-Pv 11700, left mandible with canine, p1–2 alveolus and complete p3–m2 ( Text-fig. 4b View Text-fig ). It corresponds to a small specimen, but the dentition is morphologically similar to that of other specimens. The p1 and p2 alveoli are uniradiculated and reduced, a large diastema is developed between canine–p1 and p1–p2.
NM-Pv 11695, right mandible with p2–m2 ( Text-fig. 4c View Text-fig ), mesial premolars practically unicuspidated; the p3 is unicuspid; the p4 is quite well developed, as is the main cuspid, which is higher than the m1 paraconid; a clearly separated and relatively acute distal cuspid is present. The m1 is very robust, with a short paraconid, mesial cristid somewhat inclined distally, and very pronounced. High and very bulky protoconid, with sharp mesial and distal cuspids. Quite strong metaconid, slightly displaced distally and with the lingual wall somewhat swollen. Short talonid, almost completely occupied by the hypoconid, the buccal wall of which is vertical. Low crestiform subdivided entoconid. Very weak basal cingulids. As in the m1 NM-Pv 11677, the buccal base of the hypoconid is thickened apically. The m2 has a moderately high trigonid, with the protoconid and metaconid almost the same height, although the protoconid is better developed. The protoconid presents a weak buccal widening. Very small paraconid, barely separated from the anterior cristids of the protoconid and metaconid; both join together, closing mesially the trigonid valley. Wide talonid, with a buccal hypoconid, clearly separated from the protoconid. A low, peripheral and crestiform entoconid is connected with the distal cristid of the hypoconid, clearly delimiting the talonid valley, which is flat and very wide. Basal cingulid very weak.
NM-Pv 11698, association of a right p4, m1 and m2 ( Text-fig. 4d View Text-fig 1–3 View Text-fig View Text-fig View Text-fig ). Neither the p4 nor the m1 show any differences from those described above. The m2 has a shortened talonid dominated by a very broad hypoconid. The cuspids are somewhat bunodont although they exhibit the same morphological arrangement as the previously described m2.
NM-Pv 11697, right mandible with p3–m3 ( Text-fig. 4e View Text-fig 1–2 View Text-fig View Text-fig ). It differs from NM-Pv 11695 in the greater gracility of the dentition, especially evident in m1 and m2, additionally the m2 has a relatively longer talonid. The m3 is sub-rounded and very small in size with respect to the m2.
NM-Pv 11699, left mandible with p3–m3 ( Text-fig. 4f View Text-fig 1– 3 View Text-fig View Text-fig View Text-fig ), in which the mandibular bone is only partially preserved in the mesial section (alveolus for p2 and p3–p 4 in situ), p3 unicuspid, p4 with a well-developed posterior cusp, and a thickened mesial cristid which does not form a distinct cusp. m1 robust and short, the trigonid is relatively high with respect to the talonid, the metaconid is strong. Talonid with well-developed entoconid, separated by the narrow valley of the hypoconid. The m2 is narrow with a highly developed protoconid; the talonid is short with well-developed cusps. The m3 is sub-rounded, simple, with only the protoconid clearly pronounced, a cingulum almost surrounding the entire molar.
MN-Pv 11722, association of left p4–m3 ( Text-fig. 4g View Text-fig 1–3 View Text-fig View Text-fig View Text-fig ). The p4 is tall, with the distal part widened. m1 with reduced metaconid, and talonid dominated by a strong hypoconid, the entoconid is subdivided. m2 with a narrow talonid dominated by a strong hypoconid. m3 is more compressed buccolingually than in other specimens
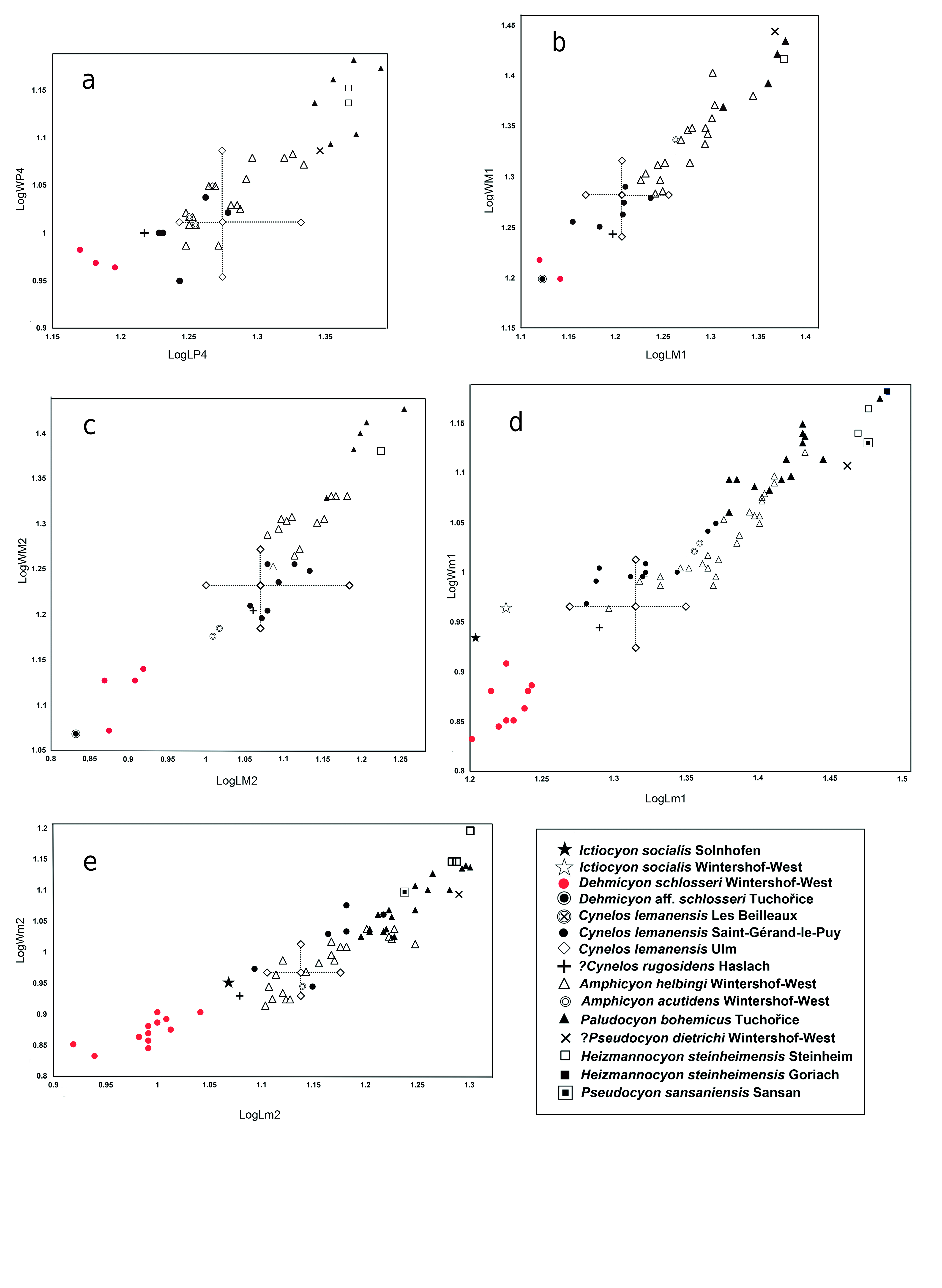
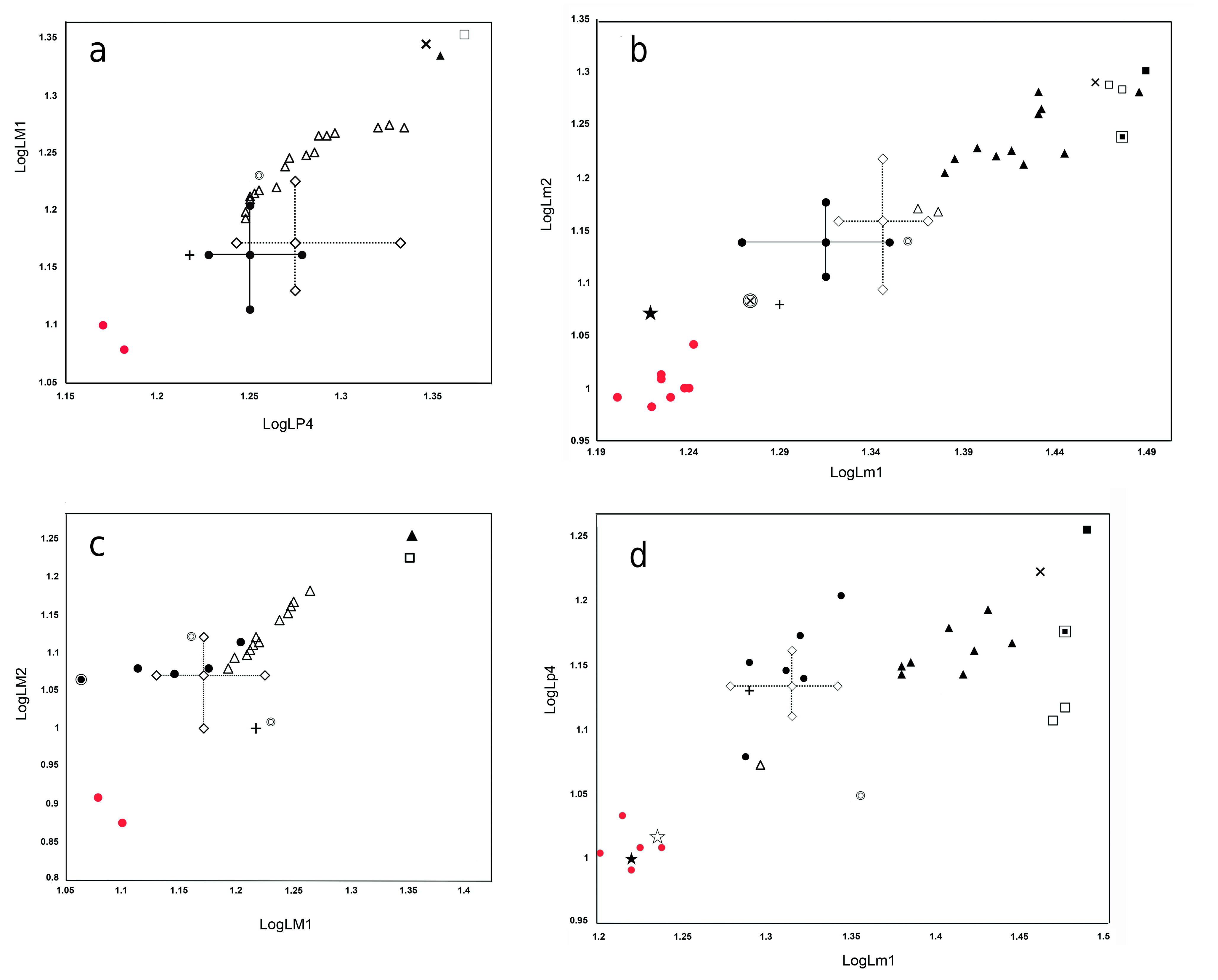
D i s c u s s i o n. Paludocyon bohemicus was originally included by Schlosser (1899) in Pseudocyon , but its systematic position has since aroused controversy (Schlosser 1891, Kuss 1965). Ginsburg (1999), in his review of the Miocene carnivorans of Europe, included it in the clade comprising Amphicyon (Heizmannocyon) bohemicus - steinheimensis with Amphicyon steinheimensis FRAAS, 1885, choosing the latter as the type species of the new subgenus. Peigné et al. (2008) considered this subgenus as a synonym of Cynelos , and even pointed out the difficulties involved in relating these two species. Hunt (2003), without establishing its validity, points out that Heizmannocyon would be closer to Cynelos than to Amphicyon , an observation most likely influenced by the retention of primitive dental characters in Cynelos , as is also the case in P. bohemicus .
As we have pointed out in the differential diagnosis of Paludocyon , it differs from Cynelos sufficiently to separate both genera. Cynelos lemanensis represents a lineage of Amphicyoninae that presents greater development of the crushing teeth, but has maintained slightly modified premolars and carnassials. Paludocyon shows a somewhat contrasting trend, characterized by a moderate increase in the size of the crushing teeth, widening of the m1 talonid, and a reduction in the size of the lower premolars. Some of these characters are found in Heizmannocyon steinheimensis but the morphology of the upper and lower molars of both genera differ to a large extent. The differences between Paludocyon and Pseudocyon sansaniensis are significant, as can be seen in our differential diagnosis. However, this comparison is limited due to the lack of well-preserved upper dentition of Pseudocyon sansaniensis, as recognized by Ginsburg (1961) and more recently by Peigné (2012). Morphologically, Heizmannocyon steinheimensis and Pseudocyon sansaniensis are similar; both share a significant reduction of the p4 as well as a narrow talonid in both the m1 and m2, but H. steinheimensis retains a large m2.
Paludocyon bohemicus is morphologically quite different from Amphicyon major from Sansan ( Ginsburg 1961); indeed, the latter shares many derived characters with Cynelos lemanensis, particularly those related to the high degree of development in the crushing dentition. Additionally, the m1 of A. major presents better development of the trigonid compared with the Paludocyon and Cynelo s species ( Ginsburg 1961). This mixture of crushing molar dentition and the high degree of development of carnassial teeth characterizes the Amphicyon species and would appear to culminate in Megamphicyon .
Paludocyon bohemicus is absent in the WintersoftWest Amphicyonidae association, although some authors considered Amphicyon dietrichi DEHM, 1950 as a very similar species, and have even proposed to synonymize both species ( Kuss 1965, Peigné et al. 2008, Hunt and Stepleton 2015). However, the revision of P. bohemicus makes it possible to discard such a close relationship. The m1 morphology suggests a greater proximity between A. dietrichi and Pseudocyon sansaniensis than with Paludocyon bohemicus . A different case could occur with the mandible from the Thenay site determined as Cynelos bohemicus by Gagnaison et al. (2012). The excellent preservation of this mandible shows that the m1 has a very wide talonid, dominated by a powerful hypoconid, morphologically the dentition is very close to that of Heizmannocyon steinheimensis, being more derived than that of Paludocyon bohemicus . Finally, Jiangzuo et al. (2020) pointed out that the first record of Cynelos cf. bohemicus , together with Cynelos cf. helbingi, is in the middle Miocene Halamagai Formation from Northwestern China. The material is scarce and alternatively to its determination as two different forms, the three specimens (M1, M2 and m2) could correspond to a single species, clearly determined by these authors as Cynelos cf. helbingi.


| TU |
Tulane University, Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Paludocyon
| Morales, Jorge, Fejfar, Oldřich, Heizmann, Elmar, Wagner, Jan, Valenciano, Alberto & Abella, Juan 2021 |
Paludocyon
| Morales & Fejfar & Heizmann & Wagner & Valenciano & Abella 2021 |
Cynelos
| , Paludocyon 2021 |
Paludocyon
| Morales & Fejfar & Heizmann & Wagner & Valenciano & Abella 2021 |
Paludocyon
| Morales & Fejfar & Heizmann & Wagner & Valenciano & Abella 2021 |
Paludocyon
| Morales & Fejfar & Heizmann & Wagner & Valenciano & Abella 2021 |
Paludocyon
| Morales & Fejfar & Heizmann & Wagner & Valenciano & Abella 2021 |
Cynelos
| , Paludocyon 2021 |
Cynelos
| , Paludocyon 2021 |
Megamphicyon
| KUSS 1965 |
Amphicyon dietrichi
| DEHM 1950 |
A. dietrichi
| DEHM 1950 |
Amphicyonidae
| TROUESSART 1885 |
Pseudocyon
| LARTET 1851 |
Amphicyon
| LARTET 1836 |