
Ameiva concolor Ruthven 1924
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3745.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:DB087293-2619-4E18-B856-8BB60409C716 |
|
DOI |
https://doi.org/10.5281/zenodo.5665429 |
|
persistent identifier |
https://treatment.plazi.org/id/970F3D7A-FFCE-C624-6683-E2ED0E8848BC |
|
treatment provided by |
Plazi |
|
scientific name |
Ameiva concolor Ruthven 1924 |
| status |
|
Figs. 8–9 View FIGURE 8 View FIGURE 9
Ameiva bifrontata concolor Ruthven, Occ. Pap. Mus. Zool. 155: 3–6. — Terra typica: Paipoy, Rio Crisnejas, 24 km from Marañòn (elevation 1067 m), province of Cajamarca, Peru. — 1924
Ameiva bifrontata concolor— Burt & Burt, Bulletin American Museum of Natural History, 61: 227–395. — 1931 Ameiva bifrontata concolor— Peters & Donoso-Barros, Smithsonian Institution Press, Washington D.C. & London: 20. — 1970 Ameiva bifrontata concolor— Peters & Donoso-Barros, Smithsonian Institution Press, Washington D.C. & London: 20. — 1986 Ameiva concolor— Harvey et al., Zootaxa, 3459: 1–156. — 2012
Diagnosis. A medium-sized Ameiva that can be distinguished from all other mainland congeners by the following combination of characters: (1) maximum known SVL of 128 mm; (2) lacking longitudinal ridge on frontal scale; (3) frontal plate divided in two subequal scales; (4) postnasals separated from prefrontals by frontonasals; (5) parietal scales usually 5; (6) median gular scales scarcely enlarged; (7) enlarged median mesoptychial scales slightly larger than largest gulars; (8) gulars posterior to the interauricular crease smaller than anterior gulars; (9) nasal suture passes centrally through nostril; (10) rostral projecting beyond the nasal suture and contacting postnasal; (11) supranasals not contacting supralabials; (12) circumorbital semicircle occasionally extending to frontal suture; (13) 31–33 enlarged ventral scales between gular and vent; (14) 10–12 longitudinal rows of ventral plates, outermost often distinctly smaller; (15) 80–93 DOM; (16) 174–196 DL; (17) postbrachials not or hardly dilated; (18) 34–41 LFT; (19) 29–35 SCF; (20) 10–21 FP; (21) vertebral region in most specimens with a trace of a pale vertebral streak.
Description and variation. Maximum known SVL in males 128 mm, maximum total length in males 404 mm (Holotype, UMMZ 59192); maximum SVL in females 85.8 mm, maximum total length in females 295.8 mm (ZFMK 91788). HL 0.26–0.32 (0.28 ± 0.02, n = 10) times SVL in both sexes; HH 0.11–0.14 (0.13 ± 0.1 n = 11) times SVL; HW 0.08–0.14 (0.11 ± 0.02, n = 10) times SVL; SL 0.59–0.80 (0.69 ± 0.08, n = 9) times the HL; ED 0.19–0.31 (0.24 ± 0.04, n = 9) times the HL; DSN 0.10–0.18 (0.13 ± 0.02, n = 9) times the HL; DNE 0.29–0.36 (0.31 ± 0.02, n = 9) times the HL; DEE 0.20–0.30 (0.23 ± 0.03, n = 9) times the HL. Tail round in cross section, tapering toward the tip; 2.25–2.67 (2.46 ± 0.16, n = 6) times SVL. Body cylindrical, AGL 0.40–0.54 (0.48 ± 0.04, n = 11) times SVL. Limbs well developed, FLL 0.33–0.44 (0.37 ± 0.04, n = 10) times SVL, HLL 0.63–0.87 (0.72 ± 0.02, n = 10) times SVL, TIL 0.15–0.21 (0.18 ± 0.02, n = 10) times SVL, foot 0.37–0.44 (0.4 ± 0.03, n = 10) times SVL.
Snout elongate, bluntly pointed; canthus rostralis distinct. Rostral in posterior part acute-angled and bordered by supranasals, in anterior part laterally stretched, projecting beyond the nasal suture and in short contact with postnasal; about as wide as high; smooth, except for a short posterolateral suture; visible from above. Supranasals almost triangular, in short medial contact, not contacting supralabials, bordered posteriorly by rhomboidal or oval frontonasal. Postnasal almost triangular, in short contact with rostral and frontonasal and in broad contact with loreal and first and second supralabials, in some specimens even in short contact with third supralabial. Oblique nasal suture passing centrally through oval nostril. Prefrontals paired and roughly pentagonal, with a medial suture longer than that between supranasals; laterally in contact with loreal and first supraocular and in some specimens in short contact with first supraciliary. Frontal plate divided transversely in two subequal scales, the suture between both scales forming a straight line; anterior frontal pentagonal, laterally in contact with first and second supraoculars, distinctly larger than posterior frontal; posterior frontal almost quadrate or rectangular, laterally in contact with second and third supraoculars. Pair of trapezoidal or irregular pentagonal frontoparietals, longer than wide and with long medial suture, fused in one specimen (ZFMK 91790); laterally separated from third supraocular by small circumorbital scales. Parietal series composed of 3–5 scales, mostly 5 including interparietal; interparietal more or less rectangular or irregularly pentagonal, higher than wide in most specimens, as wide or slightly wider than adjacent parietals, sutures with parietals straight or slightly oblique; lateral parietals irregularly shaped divided by oblique suture. Supraoculars 3–5 (mostly four) at each side, second and third largest. Circumorbital semicircle formed by 14–20 scales at left side, 29–37 scales combining both sides, bordering fourth up to middle or anterior portion of third supraoculars, occasionally extending to frontal suture and thus contacting second supraocular and separating third supraocular from frontoparietals; bordering posterior edge of last or last two supraoculars, and separating them from parietals by 2–4 rows of circumorbital scales. Laterally, all except first supraocular entirely separated from supraciliaries by a single, occasionally double row of small scales; 21–28 combining both sides. Supraciliaries 6–10 (on one side, mostly seven), first highest, middle longest, remaining ones subequal, anterior supraciliaries normally longer than those in posterior part. Loreal very large, single, in contact with postnasal, frontonasal, prefrontal, first supraciliary, all preoculars, first subocular, third and fourth supralabials and occasionally in narrow contact with second and fifth supralabial and first supraocular. Preoculars 1–2, if two, first one much smaller; second almost similar in size or slightly larger than first supraciliary, but distinctly smaller than suboculars. Suboculars three, all in contact with supralabials, normally longer than wide, second subocular longest. A curved keel reaching from preocular through first and second subocular. Postoculars 3–5. Enlarged supralabials 5–8 (mostly seven) to below center of eye; followed to commissure of mouth by 4–9 small scales. Supratemporals almost not distinguishable from surrounding scales. Temporal region with polygonal or rounded scales, smaller and almost granular centrally. External auditory meatus large, more or less round, bordered by granular scales. Tympanum recessed. All dorsal and lateral head scales juxtaposed and smooth. Mental anteriorly ellipsoid, posteriorly straight, bordered by first infralabials and postmental. Postmental single and pentagonal, in contact with first and second infralabials, followed by 7–8 pairs of enlarged chinshields. First pair (and in few specimens also second pair) in contact with infralabials and in broad medial contact. Remaining chinshields separated from infralabials by one row of small scales, and separated medially by scales of anterior gular region. Medial chin scales moderately small, slightly convex, smooth, juxtaposed, oval or polygonal, all subequal in size. Enlarged infralabials 5–7 (mostly six) to below center of eye; followed to commissure by 4–8 smaller scales. Gular region divided into two areas: anterior region with round or polygonal and flat scales in slightly oblique rows that usually remain subequal in size, delimited posteriorly by line uniting lower margin of ear openings. Posterior gular region covered by smaller polygonal or round scales in transverse rows. Mesoptychial scales slightly enlarged, in about two indistinct rows, polygonal or hexagonal, flat, smooth, and juxtaposed. Scales on nape and sides of neck slightly smaller than dorsals. Dorsals and scales on flanks granular, round, smooth, juxtaposed; slightly larger in vertebral region; 174–196 (188 ± 6.88, n = 11) DL; 80–93 (84 ± 4.69, n = 10) DOM. Ventrals large, smooth, rectangular, wider than long, in 10–12 longitudinal and 31–33 (32 ± 0.89, n = 11) transverse rows, outermost ventrals nearly as wide as those of the adjacent row or distinctly smaller; transition between ventrals and scales on flanks sharp. Preanal shield centrally with several enlarged scales, surrounded anteriorly and laterally by smaller scales; posteriorly by much smaller scales. FP 10–21 (16 ± 4.45, n = 6), in males in a continuous row along each thigh, with short gap medially; not visible in female and juvenile specimens. Each pore surrounded by four scales. Dorsal scales on tail slightly imbricate, rectangular, smaller than subcaudals, longer than wide, with a slightly oblique keel; in transverse and oblique rows, continuous with subcaudals around tail (except first few rows); 29–35 (31 ± 2.31, n = 10) SCF. Subcaudals rectangular; smaller than ventrals; wider than long close to base, longer than wide in most parts of the tail; smooth, mostly juxtaposed. Forelimbs with row of very large, smooth, slightly imbricate, almost rectangular (distinctly wider than long) antebrachial scales on anterodorsal aspect of forearms and similar but smaller brachial scales on upper arms that extend almost to insertion of forelimbs. Antebrachials and brachials usually separated by smaller scales at elbow. Dorsoposterior, posterior, and ventral aspect of arms granular, equal in size to dorsals, except for two rows of rhomboidal scales directly adjacent to antebrachials and some moderately enlarged irregular, hexagonal scales adjacent to brachials. Legs with large, smooth, imbricate scales on anterior and ventral aspects of thighs, and ventral aspect of shanks. Ventral scales on thigh, gradually becoming smaller and irregular toward pores. On ventral aspect of shanks enlarged scales arranged in two to three rows, anterior largest, dilated, and more or less trapezoidal, posterior one or two rhomboidal, decreasing in size from anterior toward posterior row. Tibiotarsal spurs not distinguishable from other scales in most specimens. Elsewhere on hindlimbs scales similar to dorsals. Supradigital scales dilated, single and smooth. 17–20 (19 ± 0.97, n = 10) LFF; 34–41 (37 ± 2.34, n = 11) LFT. Lamellae of hand transversely enlarged, convex, and single, moderately to distinctly tubercular towards base. Lamellae of outer and inner fingers continuing to wrist and only separated by few granules, tubercular and increasing in size towards it. Lamellae of toes mostly single sometimes paired; becoming distinctly tubercular towards base of second, third and fourth toe; continuing to heel on outer toe.
Coloration: No conspicuous difference in color pattern between males, females and juveniles present.
In life, color on dorsum, head and dorsal surface of limbs light to moderately brown or brownish-olive; dorsum and head with or without some small dark spots; vertebral region in most specimens with a trace of a pale vertebral streak; sides of body brownish or bluish-gray and heavily mottled with tiny dark-brown or black spots, that are often merged together, resulting in dark body sides, with in most specimens a smooth transition from the lighter vertebral region; rostral cream-colored; sides of head light reddish-brown or brownish-olive, supralabials sometimes darkly edged; tail dorsally reddish-brown or moderately brown, lightening towards tip; head and body ventrally pale yellow, cream-white or grayish-white; plates of 3 outer longitudinal rows of ventrals darkly-grayish edged, some ventrals of the outermost row even almost completely dark gray; limbs and tail ventrally cream-white; tubercular lamellae of hands and feet accentuated with brown.
In preservative, the general dorsal color is mainly grayish-brown or dark brownish-black; sides of the body are bluish–gray; the tail is grayish in anterior part fading posteriad into beige; head, body, limbs and tail ventrally pale yellowish, cream-white or grayish-white; edging of outer ventrals bluish-gray.
Distribution and natural history. This species is endemic to the dry forest of the Canyons of the Crisnejas and Marañón River in the Northern Peruvian Andes ( Fig. 3 View FIGURE 3 ) from the type locality Paipoy, Region of Cajamarca, 24 km from the Marañón (1067 m, Ruthven 1924) to Pías (Laguna), Province of Tayabamba, Region of La Libertad (1720 m, Fig. 10 View FIGURE 10 ). In addition to the locations from where we collected voucher specimens of A. concolor , we sighted this species in Vijus, Province of Pataz, Region of La Libertad, but did not collect voucher specimens. Ameiva concolor was found during daytime moving hectically on the ground in low vegetation. In Pías, the habitat was interspersed with big stones and rocks underneath which they quickly sheltered when being scared. During night time individuals were found sleeping under stones. Air temperature during the active hours of this species was between 39.3°C and 41.4°C and substrate temperature of the ground was between 33.9°C and 41.2°C. Similar to the other two Ameiva species described herein, this species was also found in the same microhabitat as Microlophus stolzmanni .
Niche comparisons. The first two axes (PC1 and PC2) of the PCA-env explain together 91.86% of the variation among the 19 bioclimatic variables in the analysis of A. aggerecusans versus A. concolor , 80.41% in the analysis of A. aggerecusans versus A. nodam and 84.59% in the analysis of A. concolor versus A. nodam . Correlation circles illustrating the contribution of each of the 19 bioclimatic variables to each PC are shown in Figure 11 View FIGURE 11 . For all three analysis the variables ‘annual mean temperature’ (BIO1), ‘temperature annual range’ (BIO7), ‘mean temperature of the wettest quarter’ (BIO8), ‘mean temperature of the warmest quarter’ (BIO10), ‘mean temperature of the coldest quarter’ (BIO11), ‘annual precipitation’ (BIO12), ‘precipitation of the wettest month’ (BIO13), and ‘max temperature of warmest month’ (BIO5) correlated negatively to PC1, whereas ‘temperature seasonality’ (BIO4), ‘mean temperature of the driest quarter’ (BIO9), and ‘min temperature of coldest month’ (BIO6) correlated positively to PC1.
A. nodam seems to use the greatest environmental space of the three species and A. concolor the smallest but this observation may be related with a higher number of species records in A. nodam .
There are a small differences in the climatic niches of A. aggerecusans and A. concolor in temperature related variables (BIO1, BIO7, BIO8, BIO10, BIO11) and precipitation related variables (BIO12, BIO13) with A. aggerecusans occupying slightly warmer and wetter regions than A. concolor ( Fig. 11 View FIGURE 11 A). The differences in PC2 between the niches of A. aggerecusans and A. concolor are only marginal. The climatic niche of A. aggerecusans is nested within the niche of A. nodam but there is a shift in the highest density of occurrences driven by precipitation and climatic seasonality (BIO14, BIO15, BIO16, BIO18, BIO19, see appendix II for a detailed description of the variables) with A. nodam occupying more humid regions with a higher climatic seasonality ( Fig. 11 View FIGURE 11 B). The differences in PC1 between the niches of A. aggerecusans and A. concolor are only marginal. The climatic niches of A. concolor and A. nodam differ in temperature related variables (BIO1, BIO7, BIO8, BIO10, BIO11) and in precipitation related variables (BIO12, BIO13) with A. nodam occupying warmer and wetter regions than A. concolor ( Fig. 11 View FIGURE 11 C). The differences in PC2 between the climatic niches of A. concolor and A. nodam are only marginal.
Pairwise comparison revealed low niche overlap values of D = 0.034 for A. aggerecusans versus A. concolor , D = 0.026 for A. aggerecusans versus A. nodam and D = 0.008 for A. concolor versus A. nodam . The hypothesis of niche equivalency was rejected for the three lineage pairs, due to significant differences between the niches of the sister taxa (niche equivalency test: P = 0.02). The results of the niche similarity are shown in Table 1 View TABLE 1 and were significant in one of the paired comparisons.
*significance value of p <0.05.
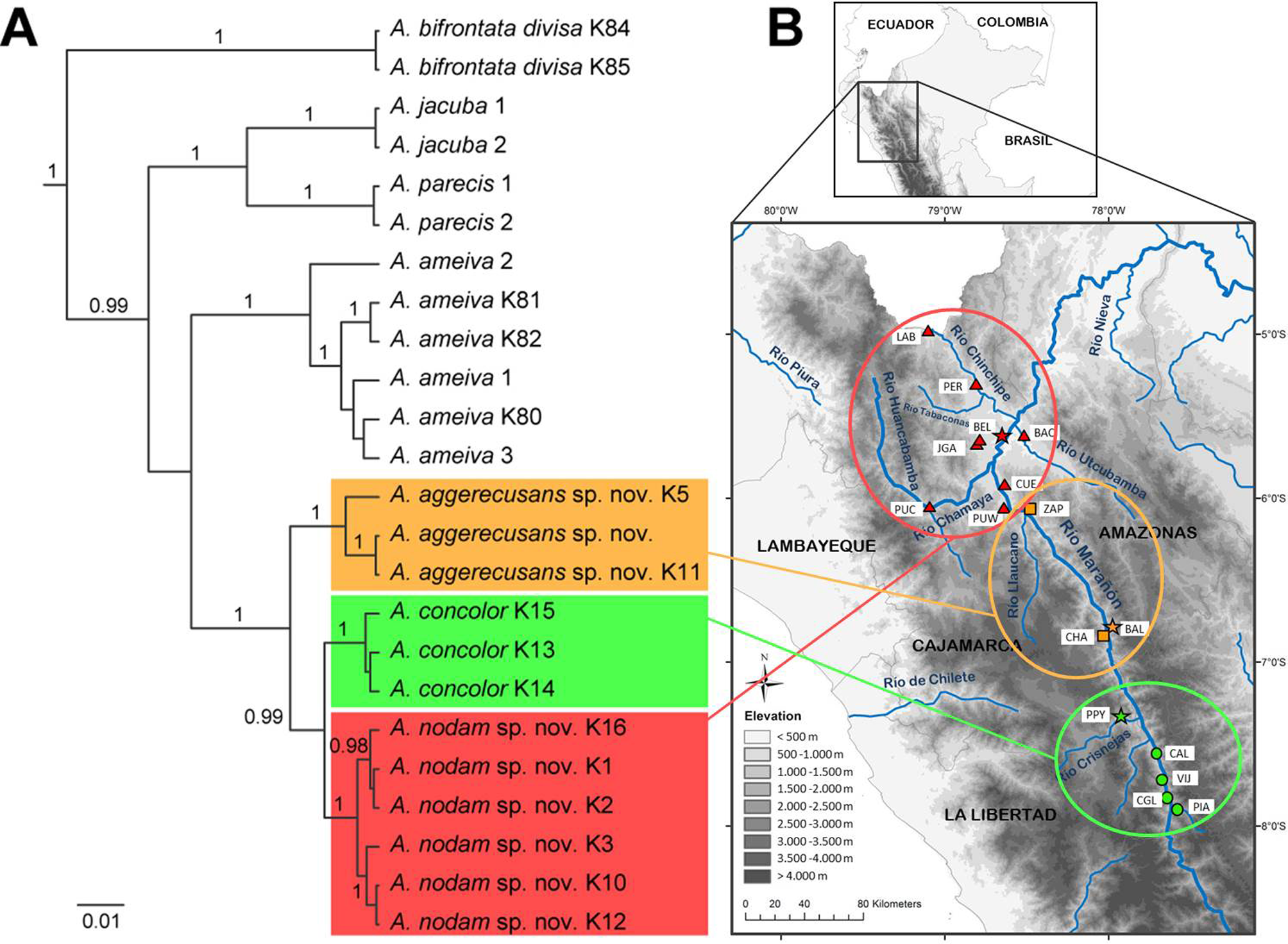




TABLE 1. Results of the pairwise niche overlap evaluation: Schoener’s D, p-values of the equivalency test and the similarity tests.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.