
Alpheus vladivostokiensis ( Vinogradov, 1950 ) Vinogradov, 1950
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4127.1.10 |
|
publication LSID |
lsid:zoobank.org:pub:0EC0211D-3392-4426-9E03-13906D799B7D |
|
DOI |
https://doi.org/10.5281/zenodo.6070163 |
|
persistent identifier |
https://treatment.plazi.org/id/3E10E143-A014-D173-FF16-FEB0FDA9F8C6 |
|
treatment provided by |
Plazi |
|
scientific name |
Alpheus vladivostokiensis ( Vinogradov, 1950 ) |
| status |
comb. nov. |
Alpheus vladivostokiensis ( Vinogradov, 1950) View in CoL comb. nov.
( Figs 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Betaeus vladivostokiensis Vinogradov 1950: 198 View in CoL , pl. 10, fig. 31; De Grave & Fransen 2011: 475 (listed as nomen dubium). Alpheus vladivostokiensis View in CoL — Pasternak 1988: 304, pl. 55, fig. 1.
Alpheus japonicus View in CoL — Balss 1914: 40 (partim); Derjugin & Kobjakowa 1935: 142 (partim?); Vinogradov 1950: 197, pl. 9, fig. 28; Marin 2013: 59, pl. 23, fig. 1 [not A. japonicus Miers, 1879 View in CoL ].
Material examined. Russia: neotype, male (cl 19.7 mm), MNHN-IU-2014-5693, Russian Far East, Primorskiy Krai, Troitza Bay (= Bukhta Troitzy), Andreevka, 42°40’4.79”N 131°7’9.24”E, off jetty, depth: 2–3 m, under rocks on mud-sand, coll. I.N. Marin, June 2011; 1 male (cl 18.0 mm), OUMNH.ZC. 2016-01-013, same collection data as for neotype; 1 ovigerous female (cl 17.4 mm), MNHN-IU-2014-5694, same collection data as for neotype; 1 ovigerous female (cl 16.3 mm), OUMNH.ZC. 2016-01-014, same collection data as for neotype; 3 males (cl 9.5– 7.5 mm), MZUSP 34289, same collection data as for neotype; 1 female (cl 12.1 mm), ZMMU Ma 608, Vladivostok, no other data, det. H. Balss as A. japonicus ; 1 male, 2 females (cl not measured, specimens in very bad condition), ZMMU Ma 1837, Krupenin, no other data, det. as A. japonicus . Japan: 1 female (cl 13.1 mm), CBM-ZC 13263, southern Hokkaido, Hakodate Bay, Hokuto City, Kattoshi, 41°44.6’N 140°36.2’E, lower intertidal (exposed at low tide), under rocks on sand and gravel, coll. S. Goshima, 22 April 2015.
Comparative material. Alpheus japonicus Miers, 1879 . Japan: 1 female (cl 10.8 mm), HUMZ-C 24, Iwate Prefecture, Miyako Bay, depth: 10–15 m, commercial round haul net, coll. T. Komai, October 1982; 1 male (cl 6.6 mm), 1 female (cl 5.1 mm), Funakoshi Bay, Iwate Prefecture, Yamada, 39°23.1’N 141°58.5’E, depth: 50 m, sand with shell fragments, dredge, coll. T. Komai, 25 May 1995; 1 female (cl 9.4 mm), MZS 70, Tanagawa, coll. L. Döderlein, 1881 [material “b” of Ortmann (1890), re-examined by Komai (1999)]; 5 males (cl 16.0–19.0 mm), 3 females (cl 16.5–18.0 mm), MZS 73, Tokyo Bay, 1880–1881 [material “a” of Ortmann (1890), re-examined by Komai (1999)]; 1 female (cl 9.6 mm), CBM-ZC 5044, Tokyo Bay, off Yokoyama, depth not recorded, 27 November 1996; 2 males (cl 8.3, 13.0 mm), 1 ovigerous female (cl 12.2 mm), CBM-ZC 10, Chiba Prefecture, Tokyo Bay, Banzu, Kisarazu, depth not recorded, scuba diving, coll. T. Furota, 10 May 1984; 1 ovigerous female (cl 10.3 mm), CBM-ZC 16, Tokyo Bay, same locality, depth not recorded, scuba diving, coll. T. Furota, 8 September 1989; 1 male (cl 13.9 mm), OUMNH.ZC. 2016-01-015, Tokyo Bay, sta. 2, depth: 30 m, beam trawl, coll. W. Doi, 18 April 2006; 1 male (cl 14.3 mm), MNHN-IU-2014-5695, same collection data as for previous specimen; 1 female (cl 12.0 mm), MNHN-IU-2014-5696, same collection data as for previous specimen; 3 males (cl 14.1– 11.3 mm), MZUSP 34290, same collection data as for previous specimen; 1 male (cl 10.6 mm), 1 female (cl 12.6 mm), CBM-ZC 8109, Seto Inland Sea, off Kurahashi Island, Hiroshima Prefecture, depth: 10–20 m, commercial trawler, coll. M. Fujiwara, 9 October 2004; 1 ovigerous female (cl 17.3 mm), Seto Inland Sea, Suonada, 33°43.0’N 131°15.0’E, depth: 18 m, TRV “Toyoshio-maru”, 2000-10 cruise, sta. 15, sledge net, coll. S. Ohtsuka, 14 July 2000; 1 female (cl 7.6 mm), Kyushu, Ohita Prefecture, Beppu Bay, 33°16.8’N 131°46.6’E, depth: 48 m, dredge, TRV “Toyoshio-maru”, 1996-5 cruise, sta. 18, coll. T. Komai, 6 June 1996. Korea: 1 female (cl 10.5 mm), CBM-ZC 5515; Kwangyang, depth: ca. 10 m, commercial trawler, coll. F. Muto, October 1999. China: 3 males (cl 13.9– 13 mm), 1 female (cl 14.6 mm), ZRC.1998.1080, Jiaozhou Bay, coll. Zhong, September 1982, det. J.Y.Liu.
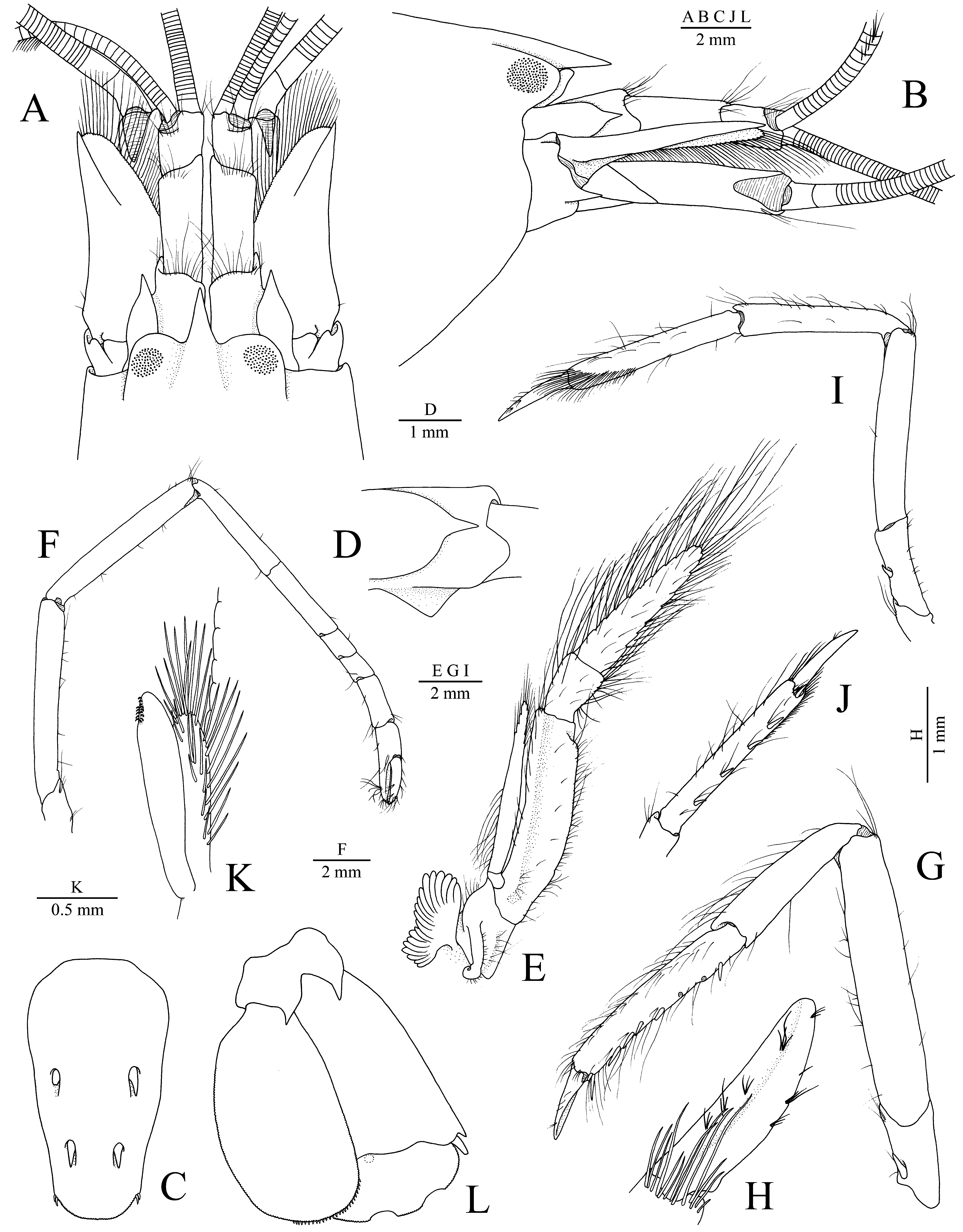
Description. Large-sized species of Alpheus edwardsii species group, cl and tl of largest specimen (neotype): 19.7 mm and 64.5 mm, respectively. Rostrum conical, acute distally; rostral carina broad, not conspicuously demarked, smoothly flattening between orbital hoods; orbital hoods moderately swollen, rounded, unarmed; adrostral furrows shallow ( Fig. 1 View FIGURE 1 A, B). Carapace with pterygostomial angle rounded ( Fig. 1 View FIGURE 1 B); cardiac notch deep. All pleonal pleura rounded marginally; sixth pleonite without articulated flap. Telson moderately broad, subrectangular, tapering posteriorly, dorsal spiniform setae well developed, situated at some distance from lateral margins, first pair anterior to telson mid-length, second pair at about 0.7–0.8 of telson length; posterior margin broadly rounded, each posterolateral angle with two small spiniform setae, lateral much shorter than mesial ( Fig. 1 View FIGURE 1 C).
Eyestalk with moderately developed, anterolaterally situated cornea; ocellar beak well developed.
Antennular peduncle with second article slightly more than twice as long as wide; stylocerite reaching or almost reaching distal margin of first article; ventromesial carina of first article with triangular and usually subacutely pointed tooth ( Fig. 1 View FIGURE 1 A, B, D). Antenna with basicerite distally ending in large ventrolateral tooth; scaphocerite relatively broad, with slightly concave lateral margin and narrow blade, latter not over-reaching very strong distolateral tooth, separated from distal portion of blade by deep cleft; carpocerite reaching slightly beyond scaphocerite and end of antennular peduncle ( Fig. 1 View FIGURE 1 A, B).
Mouthparts typical for Alpheus . Third maxilliped slender, pediform, not operculate; antepenultimate article flattened ventrolaterally; penultimate article about 1.5 times as long as high, widening distally; ultimate article about as long as antepenultimate article, gradually tapering distally, very setose; lateral plate of coxa dorsally protruding, subacute; single arthrobranch well developed ( Fig. 1 View FIGURE 1 E).
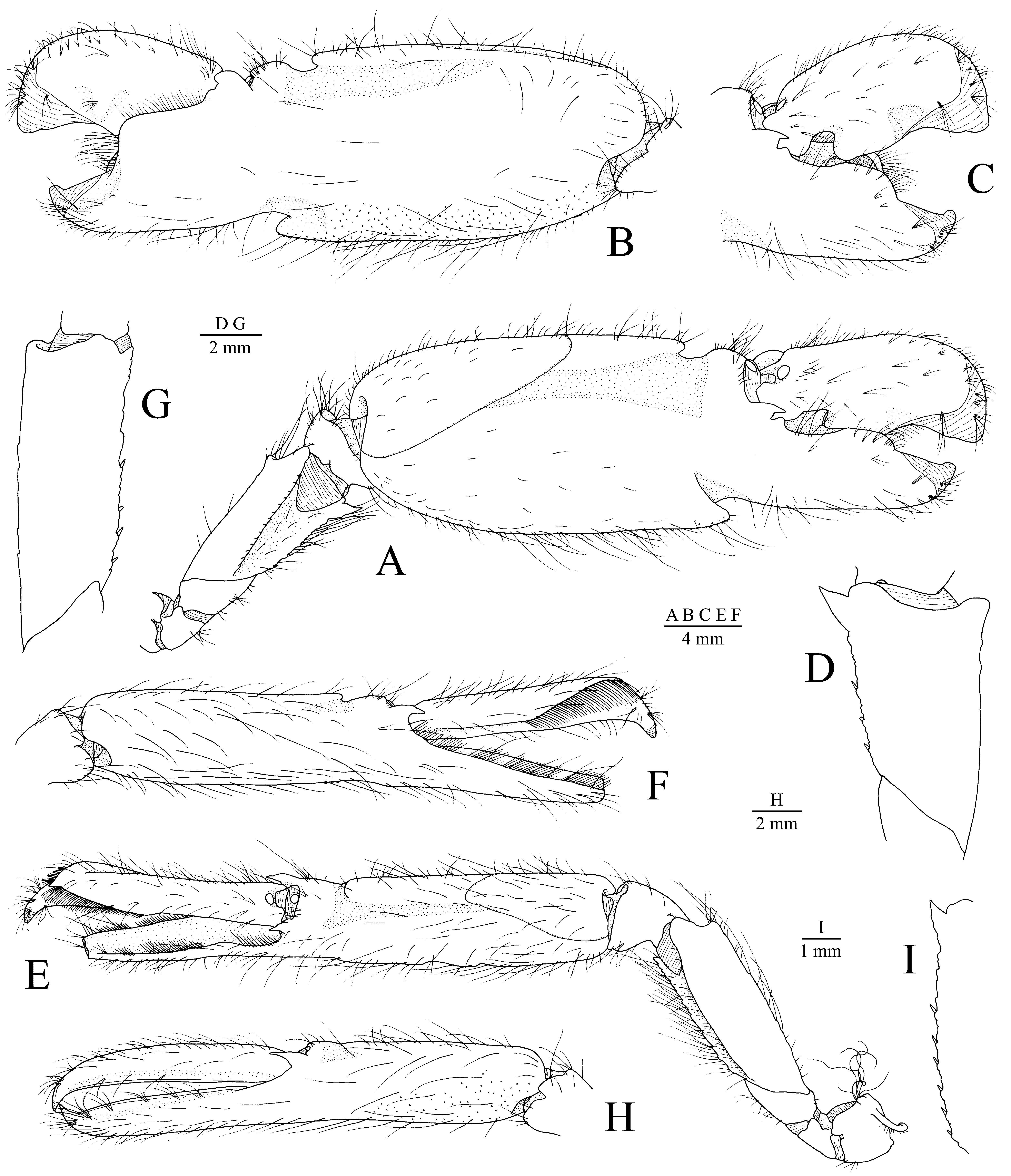
Major cheliped generally similar in both sexes, larger and with sometimes slightly different proportions in males. Ischium very short, unarmed. Merus very stout, blunt distodorsally; ventromesial margin rugose, with short spiniform setae, distally with large sharp tooth. Carpus very short, cup-shaped. Chela with palm about 1.8 times as long as fingers, distodorsal margin with broad notch extending onto lateral and mesial surfaces as shallow longitudinal grooves; dorsal shoulder distinctly overhanging distodorsal notch, rounded; ventral margin with very deep, broad notch, adjacent areas depressed on both mesial and lateral surface, ventral shoulder markedly protruding anteriorly, rounded; mesial surface of palm with small granules near ventral margin; distal margin of palm with sharp tooth laterally and acute or subacute (when not broken) tooth mesially; dactylus slightly overreaching pollex, with moderately developed, not greatly protruding plunger ( Fig. 2 View FIGURE 2 A–D).
Male minor cheliped about as long as major cheliped, however, much more slender. Ischium very short, unarmed. Merus and carpus similar to those of major cheliped, noticeably more slender. Chela with palm approximately 1.4 times as long as fingers; distodorsal margin with moderately deep notch extending onto lateral surface as shallow longitudinal groove and onto mesial surface as short triangular groove; ventral margin with shallow, sometimes barely noticeable sinus; distal margin of palm with sharp tooth on both lateral and mesial sides; ventromesial surface of palm usually with some granules; dactylus somewhat expanded, especially distally, lateral and mesial ridges of distal half furnished with balaeniceps setae; pollex also with row of setae on lateral and mesial sides ( Fig. 2 View FIGURE 2 E–G).
Female minor cheliped with ischium, merus and carpus similar to those of male minor cheliped. Chela with fingers almost as long as palm; palm with shallow distodorsal depression, not extending onto lateral and mesial surfaces, ventral margin without sinus; dactylus not expanded, without ridge and without balaeniceps setae; pollex also without balaeniceps setae ( Fig. 2 View FIGURE 2 H, I).
Second pereiopod moderately slender; merus subequal to ischium; carpus five-jointed, ratio of joints approximately equal to 5:4:1:1:2; chela as long as second carpal joint ( Fig. 1 View FIGURE 1 F). Third pereiopod relatively robust; ischium with stout ventrolateral spiniform seta; merus about five times as long as wide, unarmed; carpus about half-length of merus, with long setae on dorsal surface; propodus with numerous long setae arising especially from dorsal and ventral surfaces and row of stout spiniform setae along ventral margin, including one pair of longer distoventral spiniform setae adjacent to dactylus; dactylus about one-third of propodus length, subspatulate ( Fig. 1 View FIGURE 1 G, H). Fourth pereiopod similar to third. Fifth pereiopod shorter and more slender than third or fourth pereiopods; ischium unarmed; carpus 0.75 length of merus; propodus with well-developed setal brush distally and several stout spiniform setae along ventromesial margin; dactylus less expanded compared to that of third pereiopod ( Fig. 1 View FIGURE 1 I, J).
Male second pleopod with appendix masculina slightly shorter than appendix interna, furnished with slender spiniform setae, as illustrated ( Fig. 1 View FIGURE 1 K). Uropod with both lobes of protopod ending in acute or subacute points; exopod and endopod very broad, rounded; diaeresis of exopod slightly sinuous, lateral portion with broad rounded lobe adjacent to stout distolateral spiniform seta; distal margin of endopod with row of small spinules ( Fig. 1 View FIGURE 1 L).
Colour pattern. Overall dull olive-green or brown with some whitish areas on carapace; chelipeds greenishbrown or brown; walking legs reddish with white articulations; uropod greenish-olive proximally, bluish-grey distally; eggs olive-green ( Figs. 3 View FIGURE 3 , 4 View FIGURE 4 ; see also Pasternak 1988; Marin 2013).
Type locality. Zolotoi Rog Bay (= Bukhta Zolotoi Rog) near Vladivostok, Russia ( Vinogradov 1950); neotype from Troitza Bay (= Bukhta Troitzy), about 60 km southwest of Vladivostok.
Distribution. Northwestern Pacific: northern Sea of Japan. Presently known only from a few localities in the Russian Far East (Primorskiy Krai), e.g., Vladivostok area, Peter the Great Bay (= Zaliv Petra Velikogo), Troitza Bay (= Bukhta Troitzy), as well as from Hakodate Bay in southern Hokkaido, Japan ( Fig. 7 View FIGURE 7 ).
Ecology. Intertidal and shallow subtidal to at least 30 m; on muddy or mixed muddy-rocky bottoms; in simple burrows dug in mud or mud-sand under large rocks; typically in male-female pairs.
Remarks. Alpheus vladivostokiensis is closely related to A. japonicus ( Figs. 5 View FIGURE 5 , 6 View FIGURE 6 ), a species occurring in deeper water (below 10 m, typically 40–60 m) off the coasts of Japan, Korea, China and Taiwan ( Yokoya 1930; Liu 1955; Kim 1977; Banner & Banner 1984; Baba et al. 1986; Hayashi 1998; Anker 2001; Cha et al. 2001; Xu et al. 2015; see also Fig. 7 View FIGURE 7 ). The records from Taiwan are based on material from Dasi and Nanfang-ao fishing ports trawled off northwestern Taiwan ( Anker 2001; T.Y. Chan, pers. comm.). The two species can be distinguished from each other by a suite of morphological characters (cf. Figs. 1 View FIGURE 1 , 2 View FIGURE 2 , 5 View FIGURE 5 ). In A. vladivostokiensis , the rostral carina is slightly widening posteriorly between the orbital hoods ( Fig. 1 View FIGURE 1 A), which is not the case of A. japonicus , in which the carina remains relatively narrow ( Fig. 5 View FIGURE 5 A). The dactylus of the third and fourth pereiopods is much less expanded in A. vladivostokiensis ( Fig. 1 View FIGURE 1 H) compared to that of A. japonicus ( Fig. 5 View FIGURE 5 F). The male chelipeds in A. vladivostokiensis ( Fig. 2 View FIGURE 2 A, B, E, F) are noticeably stouter than in A. japonicus ( Fig. 5 View FIGURE 5 D, E) and also differ by the stouter, less protruding dorsal and ventral shoulders on the palm of the major cheliped, and relatively shorter, distally more expanded dactylus of the minor cheliped. In addition, A. vladivostokiensis differs from A. japonicus by the olive-green colour ( Figs. 3 View FIGURE 3 , 4 View FIGURE 4 ; see also Pasternak 1988; Marin 2013), which contrasts to the reddish colour (with a more conspicuous banding in younger individuals) of A. japonicus ( Fig. 6 View FIGURE 6 ; see also Miyake 1982; Baba et al. 1986). The ventromesial carina of the first article of the antennular peduncle exhibits some variation in both A. vladivostokiensis and A. japonicus , and may be either bluntly triangular (as in Fig. 1 View FIGURE 1 D) or with a sharp tooth (as in Fig. 5 View FIGURE 5 C).
The specimen from Vladivostok identified and reported by Balss (1914) as A. japonicus (ZMMU Ma 608) was re-examined and confirmed as A. vladivostokiensis . The listing of A. japonicus from the Sea of Japan in Derjugin & Kobjakowa (1935) is problematic as it may refer to both A. japonicus and/or A. vladisvostokiensis . As far as it can presently be ascertained, A. japonicus appears to be absent (or at least has yet not been confirmed) from the northwestern Sea of Japan (including the Russian Far East), occurring in the deeper near-shore waters of the southern Sea of Japan off the coasts of South Korea and central-southern Japan (from southern and western Hokkaido to Kyushu), as well as in the Yellow and East China Seas and the Pacific coast of Japan (from Honshu to Kyushu) ( Fig. 7 View FIGURE 7 ). Alpheus vladivostokiensis , on the other hand, seems to be confined to the shallow waters of the northern Sea of Japan, i.e. the Russian Far East and northern Japan (Hokkaido) ( Fig. 7 View FIGURE 7 ). Igarashi (1969) recorded A.
japonicus from Hokkaido on the basis of material from Oshoro Bay and Ishikari Bay, both on the Sea of Japan side of Hokkaido. The photograph of a specimen of A. japonicus (with slender chelipeds) provided by Igarashi (1969) clearly shows that his identification was correct. The listing of A. japonicus from Hokkaido in Komai et al. (1992) was based on Igarashi’s (1969) record. Thus, both A. japonicus and A. vladivostokiensis are present in southern Hokkaido, however, appear to be ecologically separated.
The somewhat confusing synonymy of A. japonicus was briefly reviewed by Banner & Banner (1984). These authors found some, possibly maturity-related variation, in the development of the balaeniceps setae on the minor cheliped. The earlier suggested synonymy of A. longimanus Spence Bate, 1888 with A. japonicus ( Ortmann 1890; De Man 1907) was confirmed, although one of Spence Bate’s specimens from India was considered to possibly belong to a different species, more closely related to A. edwardsii ( Audouin, 1826) . Part of the Japanese material identified as A. haanii Ortmann, 1890 by Yokoya (1939) was tentatively re-assigned to A. japonicus , whereas A. haanii was considered as a possible synonym of A. edwardsii . However, since the latter species is clearly a large species complex (Anker & De Grave, in press), the taxonomic status of A. haanii remains unsettled. Ortmann (1890) based the description of A. haanii on two lots, each with three specimens, from two different Japanese localities, one from Tokyo Bay and one from Kagoshima. Komai (1999) re-examined Ortmann’s Kagoshima material (MZS 69) and found that one of the three specimens in this lot represented A. bisincisus De Haan, 1844 . The other two specimens could be identified only as an unidentifiable species of the A. edwardsii species group, morphologically similar to A. lobidens De Haan, 1844 . On the other side, we concur with the conclusion of Ortmann (1890), De Man (1907) and Banner & Banner (1984) on the status of A. longimanus as a junior synonym of A. japonicus s.str. With A. haanii ruled out and with no other available name for a taxon morphologically similar to A. japonicus , the application of the name A. vladivostokiensis for A. japonicus sensu Vinogradov (1950) and Marin (2013) is now well supported.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Caridea |
|
Family |
|
|
Genus |
Alpheus vladivostokiensis ( Vinogradov, 1950 )
| Anker, Arthur, Marin, Ivan N., Pachelle, Paulo P. G. & Komai, Tomoyuki 2016 |
Betaeus vladivostokiensis
| De 2011: 475 |
| Pasternak 1988: 304 |
| Vinogradov 1950: 198 |
Alpheus japonicus
| Marin 2013: 59 |
| Vinogradov 1950: 197 |
| Derjugin 1935: 142 |
| Balss 1914: 40 |