
Alpheus echiurophilus, Anker, Arthur, Komai, Tomoyuki & Marin, Ivan N., 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3914.4.4 |
|
publication LSID |
lsid:zoobank.org:pub:7B4A8A8B-DABD-4DF4-92B0-C9F61110C10B |
|
DOI |
https://doi.org/10.5281/zenodo.6097840 |
|
persistent identifier |
https://treatment.plazi.org/id/03EE87CF-B20F-FF82-20F5-1E412FF2FEE8 |
|
treatment provided by |
Plazi |
|
scientific name |
Alpheus echiurophilus |
| status |
sp. nov. |
Alpheus echiurophilus View in CoL sp. nov.
Figs. 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , 7A View FIGURE 7. A , B, 8A, B
Type material. Japan, Ryukyu Islands. Holotype: male (cl 11.6 mm), CBM-ZC 12590, Okinawa Island, Naha, Ohmine, intertidal sand flat, yabby pump, burrow of non-identified echiuran ( Thalassematidae ), coll. T. Komai, 25.IV.2014. Paratypes: 1 male (cl 8.8 mm), 1 female (cl 10.9 mm), 3 ovigerous females (cl 10.1–11.0 mm), CBM- ZC 12591, same collection data as for holotype; 1 male (cl 7.8 mm), 1 female (cl 7.7 mm), CBM-ZC 12592, Okinawa Island, Sesoko Islet, intertidal, mixed sand-gravel substrate, under rock, burrow of echiuran (possibly Ochetostoma erythrogrammon ), coll. T. Komai, 25.V.2014; 1 male (cl 13.2 mm), CBM-ZC 12593, Yaeyama Islands, Ishigaki Island, Kabira Bay, intertidal sand flat, yabby pump, coll. T. Komai, 10.XII.2008.
Additional material. Vietnam, Nha Trang Bay. 1 male (cl 7.1 mm), OUMNH.ZC. 2014-01-0061, southeast of Tre Island, Dam Bay, 12°12’19.53”N 109°18’11.12”E, mud-sand flat off mangrove, depth: 0.5 m, yabby pump, burrow of echiuran Ikedosoma sp. (host deposited under CBM-ZW 1100) coll. I. Marin & S. Sinelnikov, 06.IV.2014.
Alpheus cf. echiurophilus sp. nov. Madagascar, Nosy-Bé. 1 male (cl 7.9 mm), FLMNH UF 14445, east of Hellville, near CNRO station, 13.4069’ S 48.2917 ’E mud-sand flat off mangrove, depth: 0.2–0.5 m, yabby pump, burrow of echiuran Ochetostoma (?) zanzibarense (host deposited under FLMNH UF 3716), coll. A. Anker, 18.V.2008 (see Remarks).
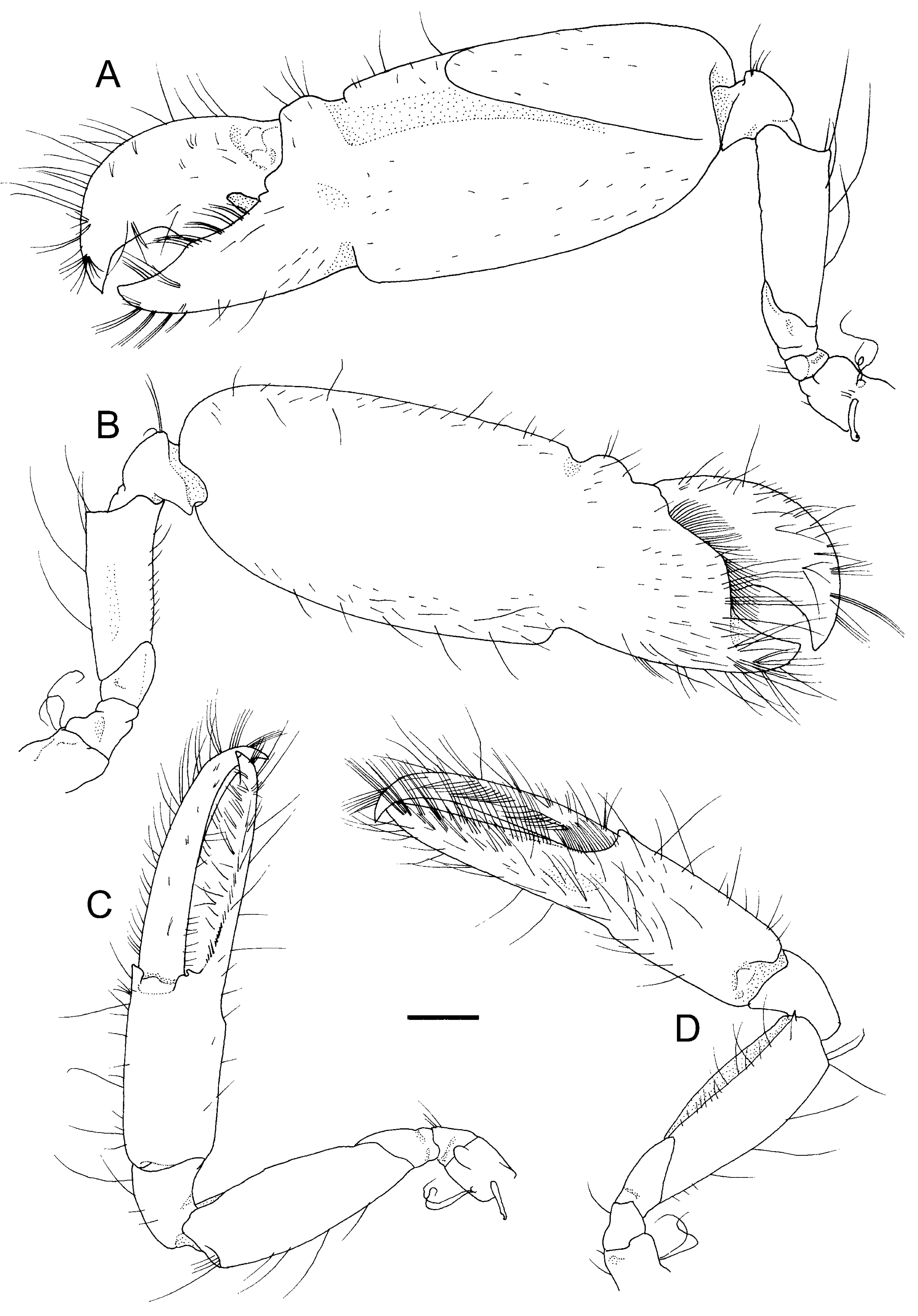
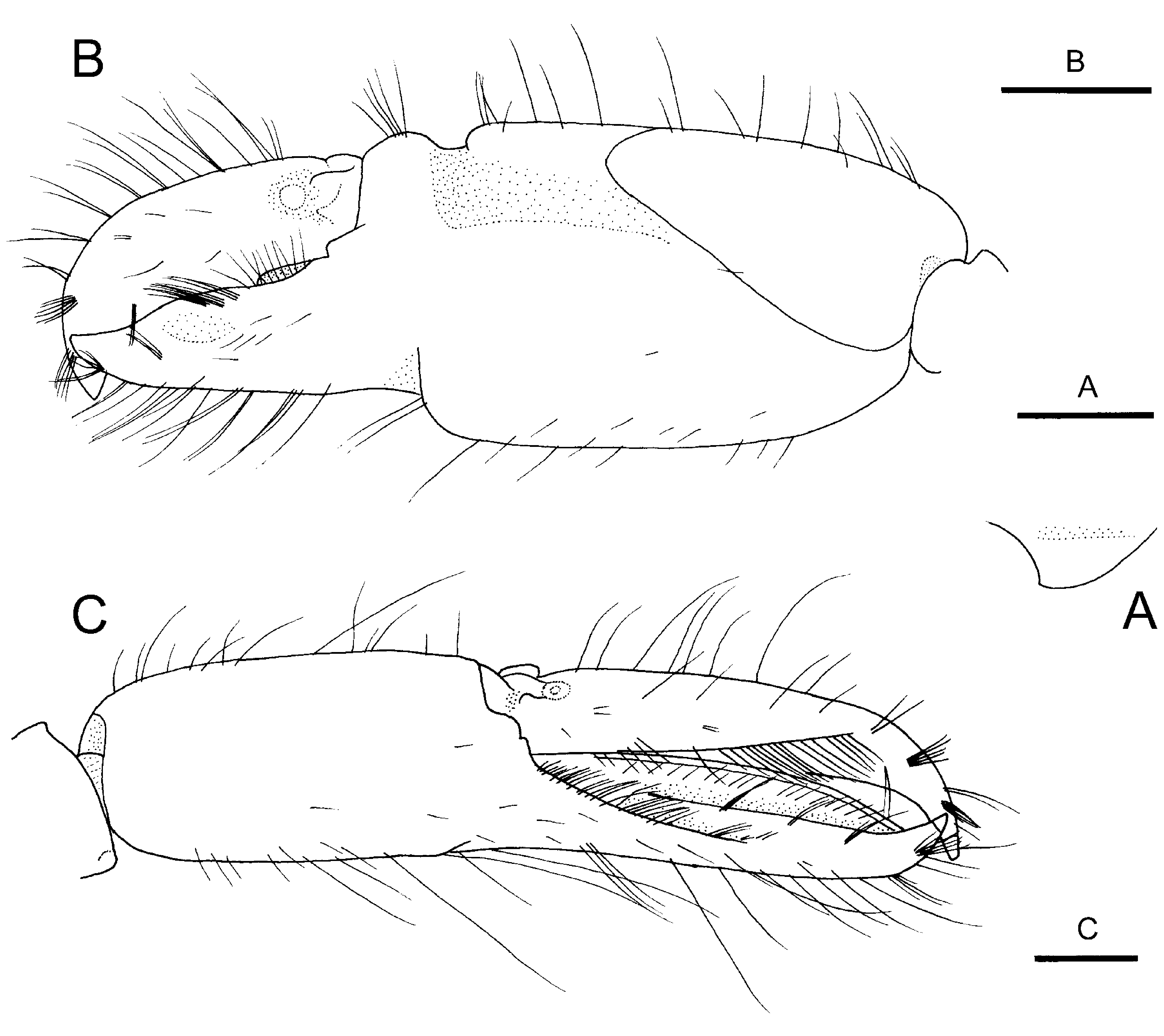
Description. Carapace ( Fig. 1 View FIGURE 1 A, B) smooth, glabrous, moderately compressed laterally; rostrum ( Fig. 1 View FIGURE 1 A, B) short, slender, directed forwards in males, usually slightly descending in females, distally acuminate, reaching midlength of first article of antennular peduncle; postrostral carina ( Fig. 1 View FIGURE 1 A) broad, rounded transversely, slightly broadened posteriorly to level of posterior margin of corneas; adrostral grooves shallow, not abruptly delimited; orbito-rostral process feebly developed; orbital hoods ( Fig. 1 View FIGURE 1 A, B) moderately inflated, distally rounded, unarmed; anterior margin between rostrum and orbital hoods nearly straight; pterygostomial angle ( Fig. 1 View FIGURE 1 B) broadly rounded; cardiac notch deep.
Pleon without distinctive features; pleura broadly rounded marginally; sixth pleonite without articulated flap, preanal area not protruding as tooth. Telson ( Fig. 1 View FIGURE 1 C) relatively broad, subrectangular, narrowing posteriorly; dorsal surface with two pairs of moderately strong spiniform setae, situated at some distance from lateral margins, first pair slightly anterior to mid-length, second pair at about 0.7 telson length; posterior margin ( Fig. 1 View FIGURE 1 D) broadly rounded, with two pairs of posterolateral spiniform setae, mesial much longer than lateral, and with row of spinules between mesial spiniform setae.
Eyes ( Fig. 1 View FIGURE 1 A, B) completely concealed in dorsal and lateral views, partly visible in frontal view, with small low tubercle on anteromesial margin; cornea well developed. Ocellar beak not conspicuously protruding between eyes.
Antennular peduncle ( Fig. 1 View FIGURE 1 A, B) moderately slender; stylocerite with acute tip, not reaching distal margin of first article; first article with ventromesial carina bearing broadly subtriangular tooth terminating in anteriorly directed, acute point ( Fig. 1 View FIGURE 1 E); second article approximately 3.0 times as long as broad; lateral flagellum with numerous tufts of aesthetascs on approximately 11th to 20th joints; secondary ramus rudimentary.
Antenna ( Fig. 1 View FIGURE 1 A, B) with basicerite bearing moderately small, slender, ventrolateral tooth; carpocerite stout, reaching far beyond distolateral tooth of scaphocerite; scaphocerite ( Fig. 1 View FIGURE 1 F) reaching distal end of antennular peduncle by distal margin of lamella, with faintly sinuous lateral margin and strong distolateral tooth, latter reaching far beyond lamella; cleft between lamella and distolateral tooth deep; lamella narrowly rounded distally.
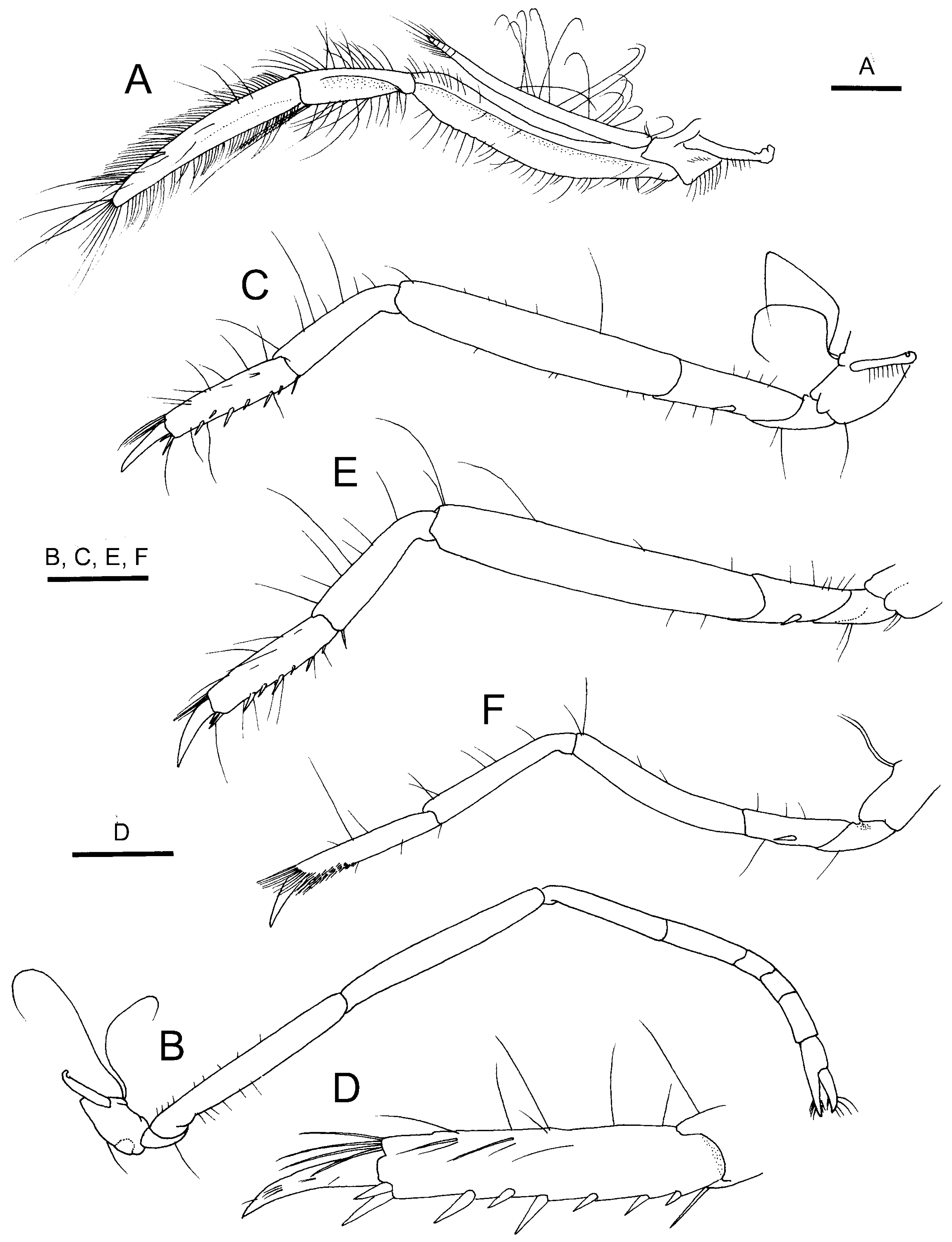
Mouthparts typical for Alpheus . Third maxilliped ( Fig. 6 View FIGURE 6 A) moderately stout; lateral plate on coxa extending to base of exopod, with subacute point; antepenultimate article subtriangular in cross-section, with distinctly keeled dorsolateral margin and sharply carinate ventral margin; penultimate article flattened dorsoventrally, about twice as long as wide in dorsal or ventral view; ultimate article distally tapering, trigonal in cross section, tip blunt with tufts of long setae.
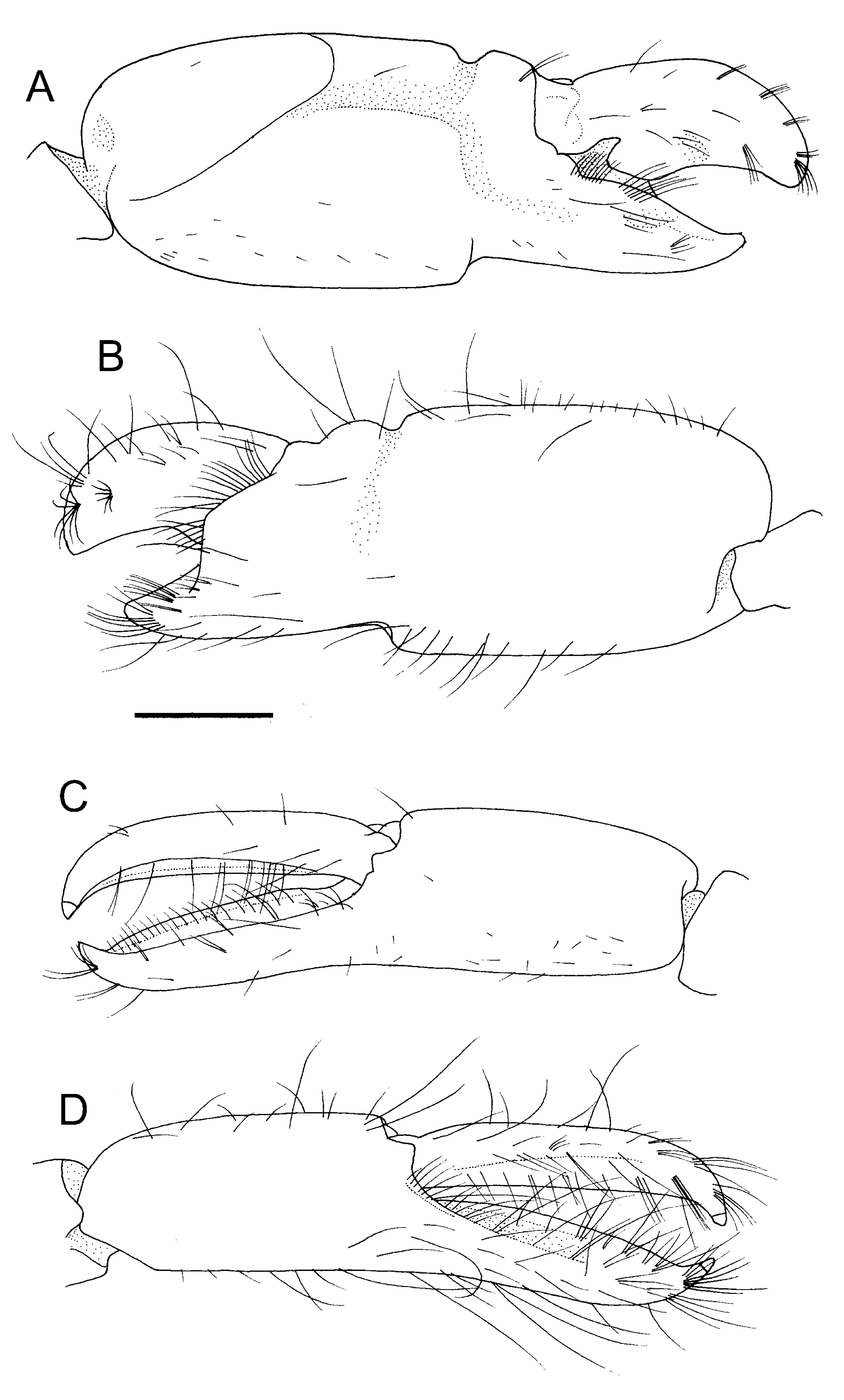
Male major cheliped ( Fig. 2 View FIGURE 2 A, B) moderately stout, much larger than minor cheliped. Ischium short, unarmed. Merus moderately stout, broadened distally, trigonal in cross-section; dorsal margin nearly straight, distodorsal margin slightly produced; ventrolateral margin faintly crenulate; ventromesial margin straight or faintly sinuous, nearly smooth, with row of short and long, unarmed ( Fig. 5 View FIGURE 5 A) or armed with 1 short spiniform seta ( Fig. 5 View FIGURE 5 C), terminating distally in small tooth. Carpus very short, cup-shaped. Chela moderately compressed laterally, slightly elongate; fingers slightly less than half-length of palm. Palm about 2.0 times as long as wide, dorsal and ventral margins with broad transverse grooves; dorsal groove only slightly extending onto mesial surface, but not continuing posteriorly, extending as shallow depression onto lateral surface continuing posteriorly along linea impressa; dorsal shoulder rounded, gently sloping, not overhanging groove; ventral groove broad, relatively shallow; ventral shoulder rounded, not protruding; linea impressa well marked; adhesive disks small. Pollex less than half-length of palm, with broad (about 120°) angle on dorsal margin, latter with fairly dense row of setae; mesial surface slightly depressed. Dactylus reaching slightly beyond pollex, without transverse ridge near base of plunger; plunger ( Fig. 3 View FIGURE 3 A) moderately long, stout. Female major cheliped ( Fig. 4 View FIGURE 4 A, B) generally similar to male major cheliped, with proportionally smaller chela; ventromesial margin of merus unarmed or armed with 1 or 2 minute spiniform setae ( Fig. 5 View FIGURE 5 E); palm about 1.8 times as long as wide.
Male minor cheliped ( Figs. 2 View FIGURE 2 C, D; 3B, C) distinctly shorter than major cheliped. Ischium similar to that of major cheliped. Merus with dorsal margin slightly convex, distodorsal margin not produced; ventrolateral margin faintly convex; ventromesial margin entire or faintly crenulate, bearing row of short to long setae, unarmed or armed with 1 or 2 minute spiniform setae ( Fig. 5 View FIGURE 5 B, D), terminating distally in small tooth. Carpus short, cupshaped, somewhat longer than that of major cheliped. Chela subcylindrical, with very shallow depression on ventral margin at pollex base. Palm about 2.2 times as long as wide, with scattered long setae on dorsal margin and lateral surface proximal to base of pollex; distomesial process near articulation with dactylus bluntly triangular. Fingers 1.2–1.3 times as long as palm, with strongly curved, crossing tips. Pollex with lateral surface bearing blunt longitudinal ridge furnished with minute spiniform setae proximally and tufts of long setae distally, surface near cutting edge excavated with two sparse rows of long and short setae; mesial surface with short longitudinal row of densely inserted setae on dorsal margin, adjacent to base of dactylus, and numerous scattered tufts of long setae; cuttinge edge sharp, blade-like. Dactylus not expanded, with lateral surface ( Fig. 3 View FIGURE 3 C) sharply carinate, bearing row of stiff setae adjacent to cutting edge; surface along cutting margin depressed, flattened, with numerous setae; mesial surface ( Fig. 3 View FIGURE 3 B) with weak, obliquely longitudinal ridge extending from dactylar base to distal 0.4 length of dactylus, longitudinal ridge furnished with row of stiff setae (proximal setae short, bristle-like, becoming gradually longer distally) and scattered long stiff setae; cutting edge sharp, blade-like.
Female minor cheliped ( Fig. 4 View FIGURE 4 C, D) generally similar to that of male, but relatively more stout; ventromesial margin of merus faintly crenulate or almost smooth, unarmed or with 1 or 2 minute, widely spaced, spiniform setae ( Fig. 5 View FIGURE 5 F); palm about 1.8 times as long as wide, without shallow depression on ventral margin near pollex base; pollex with short longitudinal ridge bearing short stiff setae proximally on mesial surface, as well as scattered tufts and individual short or long setae on mesial and lateral surfaces; dactylus without longitudinal ridge on mesial side, with scattered setal tufts or individual setae.
Second pereiopod ( Fig. 6 View FIGURE 6 B) slender, overreaching distal end of antennular peduncle by 0.9 length of carpus; ischium subequal in length to merus; carpus divided into five articles, with approximate ratio 1: 0.8: 0.3: 0.3: 0.5; chela subequal in length to second carpal article, fingers slightly longer than palm. Third and fourth pereiopods similar in shape, length and armature ( Fig. 6 View FIGURE 6 C, E). Third pereiopod ( Fig. 6 View FIGURE 6 C, D) moderately slender; ischium armed with small ventrolateral spiniform seta; merus unarmed, about 5.4 times as long as wide; carpus distinctly slenderer than merus, with one stout seta at distoventral angle; propodus 1.1 times as long as carpus, armed with four to seven spiniform setae of unequal size on ventral margin, arranged in double row, and with pair of spiniform setae at distoventral angle adjacent to dactylus; dactylus simple, subconical, with acute tip, slightly curved, 0.4–0.5 length of propodus. Fifth pereiopod ( Fig. 6 View FIGURE 6 F) shorter and slenderer than third and fourth pereiopods; ischium armed with small ventrolateral spiniform seta; propodus with grooming brush consisting of six or seven transverse rows of stiff setulose setae on distal 0.4 length of article, one small spiniform seta present at distoventral angle (usually concealed by grooming setae, not visible in lateral view); dactylus similar to that of third or fourth pereiopods.
Male second pleopod with appendix masculina ( Fig. 1 View FIGURE 1 G) distinctly longer than appendix interna, furnished with slender setae, increasing in length distally, on dorsal and distal-most surface.
Uropod ( Fig. 1 View FIGURE 1 H, I) with protopod armed with 2 large acute distal teeth; exopod with slightly convex lateral margin, terminating in small distolateral tooth, with adjacent movable spiniform seta; diaeresis faintly trilobate; distal margin with row of minute spinules dorsal to main setal fringe; endopod with scattered short to long setae proximally; distal margin with row of minute spinules dorsal to main setal fringe.
Gill/exopod formula typical for Alpheus .
Variation. Within the Japanese material of A. echiurophilus sp. nov., morphological variation exists in the number of spiniform setae (4–7) on the third and fourth pereiopods; the presence or absence of minute spiniform setae on the ventromesial margin of the cheliped meri, as illustrated ( Fig. 5 View FIGURE 5 ); the size and proportions of the major chela; and the setation of the minor cheliped. The latter two variable features are subject of sexual dimorphism, typical for many species of the genus Alpheus . The Vietnamese male specimen agrees with the holotype in most diagnostic features, except for the much less developed setae on the mesial face of the minor cheliped fingers, possibly due to its younger age (cl 7.1 mm vs. 11.6 mm in the holotype). The Madagascan male specimen, here preliminarily assigned to A. cf. echiurophilus sp. nov., presents some more substantial differences to the type series, which are discussed below (see Remarks).
Colouration. Carapace nearly colorless with pale orange tinge; pleon with pale reddish or orange chromatophores forming diffuse transverse bands across posterior portion of each somite; antennular and antennal peduncles reddish or pale orange; chelipeds with more intense reddish or orange areas, especially on distal portion of palm and on fingers; second to fifth pereiopods and pleopods colourless; tail fan with some pale orange markings; eggs yolk-yellow ( Figs. 7A View FIGURE 7. A , B; 8A, B, D).
Type locality. Ohmine, near Naha International Airport, Okinawa Island, Ryukyu Islands, southern Japan.
Distribution. Southern Japan (Ryukyu Islands: Okinawa and Ishigaki Islands) and Vietnam (Nha Trang Bay); presence in Madagascar (Nosy-Bé) requires confirmation (see below for comments).
Etymology. The specific epithet echiurophilus is a combination of the Latin words Echiura (spoon worms) and philus (originally from Greek philos), indicating love of or fondness for, referring to the new species symbiotic life style in echiuran burrows; used as an adjective.
Ecology. All specimens from Ohmine, Okinawa Island, were found in burrows of an unidentified echiuran from the thalassematid subfamily Thalassematinae , on intertidal sand flats ( Fig. 7 View FIGURE 7. A C, D). The single individual from Kabira Bay, Ishigaki Island, was also collected from an intertidal burrow; its host was not collected. The pair from Sesoko Islet, Okinawa Island, was found on sand-gravel substrate under a large rock, together with its echiuran host tentatively identified as Ochetostoma erythrogrammon Leuckart & Rüppel, 1828 (Thalassematidae) [host identification based on Nomura (2000), requires confirmation]. Another snapping shrimp, Alpheus barbatus Coutière, 1897 , a species not closely related to A. echiurophilus sp. nov., but associated with the same echiuran host, was also collected at Sesoko Islet (T. Komai, pers. obs.). The Nha Trang and Nosy-Bé specimens were both collected on sand-mud bottoms near mangroves, in very shallow water (0.2–0.5 m), from burrows of thalassematine echiurans identified as Ikedosoma sp. ( Fig. 8 View FIGURE 8 C) and Ochetostoma zanzibarense Stephen & Robertson, 1952 ( Fig. 8 View FIGURE 8 E) ( Thalassematidae ), respectively. Thus, A. echiurophilus sp. nov. appears to be an obligate associate of burrowing echiurans of the thalassematid subfamily Thalassematinae , in the intertidal and shallow subtidal habitats.
Remarks. Alpheus echiurophilus sp. nov. plainly belongs to the A. leviusculus Dana, 1852 species group, which was recently separated from the heterogeneous A. edwardsii ( Audouin, 1826) species group by Anker et al. (2009). The A. leviusculus group is similar to the A. edwardsii group in the major chela having well-developed dorsal and ventral transverse grooves. However, in the A. leviusculus group, the dorsal groove only slightly extends onto the mesial surface, not continuing proximally, whereas in the A. edwardsii group, the dorsal groove extends largely onto the mesial face of the palm, continuing proximally. Within the A. leviusculus group, A. echiurophilus sp. nov. is morphologically closest only to a few species, viz. the Indo-West Pacific A. leviusculus Dana, 1852 (including A. bastardi Coutière, 1898 according to Banner & Banner 1964, 1982, but see below), A. hululensis Coutière, 1905 , A. ladronis Banner, 1956 , and the western Atlantic A. zimmermani Anker, 2007 . All other species of the A. leviusculus group (see Anker et al. 2009) appear to be more distantly related to A. echiurophilus sp. nov., e.g., in the shape and proportions of the major and minor chelipeds, or armature and proportions of the third to fifth pereiopods.
Alpheus echiurophilus View in CoL sp. nov. can be separated from A. leviusculus View in CoL by numerous characters, including the distinctly longer second article of the antennular peduncle (three times as long as wide in the new species vs. less than twice in A. leviusculus View in CoL ); the dactylus of the major chela with a noticeably shorter plunger; the male minor chela with a shallow ventral sinus and longer fingers, latter also with a different setation; and the presence of a stout spiniform seta on the ischium of the third and fourth pereiopods (absent in A. leviusculus View in CoL ) (cf. Banner & Banner 1982: fig. 77). In addition, the colour patterns of A. echiurophilus View in CoL sp. nov. and A. leviusculus View in CoL are quite different: the former species has diffuse pale orange transverse bands on each pleonite, orange chelae and yellow eggs ( Figs. 7A View FIGURE 7. A , B, 8A, B, D), whereas the latter species has narrow green transverse bands on each pleonite, greenish chelae and green eggs ( Banner & Banner 1982).
Banner & Banner (1964, 1982) treated Alpheus Bouvieri View in CoL var. bastardi Coutière, 1898 (= A. Bastardi View in CoL in Coutière 1905) as a junior synonym of A. leviusculus View in CoL . However, Coutière (1899) described the colour of A. Bouvieri View in CoL var. Bastardi from Djibouti as uniformly orange-red, which strikingly differs from the description of the colour pattern of A. leviusculus View in CoL in Banner & Banner (1982), suggesting that A. bastardi View in CoL may well be a valid species (A. Anker, in study). In any case, A. echiurophilus View in CoL sp. nov. can be distinguished from A. bastardi View in CoL using the same morphological criteria as for A. leviusculus View in CoL .
The new species differs from the poorly described A. hululensis View in CoL by the postrostral area with a much broader, posteriorly widening carina (vs. with a relatively sharp carina in A. hululensis View in CoL ); the stylocerite not reaching the distal margin of the first article of the antennular peduncle (vs. distinctly overreaching this margin in A. hululensis View in CoL ); and the ratio of the carpal articles in the second pereiopod (first article at least 1.5 times longer than second in the new species vs. only 1.1 times in A. hululensis View in CoL ) (cf. Coutière 1905: fig. 46).
Alpheus echiurophilus View in CoL sp. nov. can be most easily distinguished from A. ladronis View in CoL by the longer second article of the antennular peduncle (three times as long as wide in the new species vs. at most twice as long as wide in A. ladronis View in CoL ); the male minor chela with dense setal rows on the fingers (absent in A. ladronis View in CoL ); and the presence of a robust spiniform seta on the ischium of the third and fourth pereiopods (absent in A. ladronis View in CoL ) (cf. Banner 1956: fig. 20).
Finally, A. echiurophilus View in CoL sp. nov. differs from A. zimmermani View in CoL by the absence of a long postrostral carina (extending beyond carapace mid-length in A. zimmermani View in CoL ); the longer distolateral tooth of the scacphocerite (reaching far beyond lamella in the new species vs. only slightly beyond in A. zimmermani View in CoL ); the normally developed cornea (reduced to a lateral bean-shaped area in A. zimmermani View in CoL ); the presence of setal rows on the fingers of the minor chela of adult males (absent in A. zimmermani View in CoL ); and the overall pale-orange colouration, with diffuse transverse bands on the abdomen, contrasting with a striking, diagnostic, red-and-white colour pattern of A. zimmermani View in CoL (cf. Anker 2007: figs. 1–3).
Alpheus echiurophilus View in CoL sp. nov. is the first species of the A. leviusculus View in CoL group known to be associated with echiuran burrows. All other echiuran-associated species of Alpheus View in CoL belong to the A. brevirostris ( Olivier, 1811) View in CoL group, and within this group to the morphologically well-defined A. barbatus Coutière, 1897 View in CoL species complex, presently containing five species ( Anker & Dworschak 2004; Anker et al. 2007). Thus, symbiosis with echiurans evolved independently twice within Alpheus View in CoL , once in the A. leviusculus View in CoL group (in A. echiurophilus View in CoL sp. nov.) and once in the A. brevirostris View in CoL group (in the common ancestor of the A. barbatus View in CoL complex). Interestingly, A. echiurophilus View in CoL sp. nov. presents only one morphological feature that may be interpreted as convergent with some members of the A. barbatus View in CoL complex. This feature is the general shape of the minor chela, with elongated fingers bearing rows of numerous elongate setae, especially in males. In the A. barbatus View in CoL complex, the development of setal rows on the minor chela is variable, being minimal in A. ribeiroae Anker & Dworschak, 2004 View in CoL and extreme in A. barbatus View in CoL ( Banner & Banner 1982; Anker & Dworschak 2004; Anker et al. 2007).
The male specimen from Nosy-Bé, Madagascar (FLMNH UF 14445), differs from the Japanese and Vietnamese male specimens in several aspects, e.g., the ventromesial carina of the first article of the antennular peduncle having a blunt tooth ( Fig. 9A View FIGURE 9. A ); the dactylus of the major cheliped being slightly longer (0.6 times as long as the palm vs. 0.5 times as long in the holotype; Fig. 9 View FIGURE 9. A B), and the minor cheliped with a distinct balaeniceps ridge on the mesial surface of the dactylus ( Fig. 9 View FIGURE 9. A C). In view of these differences and the presence of numerous species complexes in the genus Alpheus View in CoL , as well as the significant geographical distance between Japan / Vietnam and Madagascar, the Nosy-Bé specimen is preliminarily referred to Alpheus cf. echiurophilus View in CoL sp. nov. Additional material from the southwestern Indian Ocean and/or a genetic comparison (e.g., barcoding segment of COI gene) between available materials of A. echiurophilus View in CoL and A. cf. echiurophilus View in CoL are needed to elucidate the taxonomic identity of the latter taxon.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Alpheus echiurophilus
| Anker, Arthur, Komai, Tomoyuki & Marin, Ivan N. 2015 |
A. ribeiroae
| Anker & Dworschak 2004 |
Ochetostoma zanzibarense
| Stephen & Robertson 1952 |
var. bastardi Coutière, 1898
| Coutiere 1898 |
A. barbatus Coutière, 1897
| Coutiere 1897 |
A. brevirostris (
| Olivier 1811 |