
Alloionema californicum, Nermuť, Jiří, Půža, Vladimír, Mráček, Zdeněk & Lewis, Edwin, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4184.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:606F956E-56BC-46A7-AF48-F77E189713E3 |
|
DOI |
https://doi.org/10.5281/zenodo.5622471 |
|
persistent identifier |
https://treatment.plazi.org/id/305487B4-0C1B-2D06-FF22-11B88B14FDA7 |
|
treatment provided by |
Plazi |
|
scientific name |
Alloionema californicum |
| status |
sp. nov. |
Alloionema californicum n. sp.
( Figs 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Measurements (JU2015 strain). See Table 1 View TABLE 1 .
Description (based on JU2015 strain)
Adults. Six fused apparent lips surrounding triangular mouth opening, each lip with a labial papilla on top. Two lateral lips with amphidial opening. Cuticle smooth, annulated, annuli ca 1 µm wide, lateral fields not developed. Stoma ca 2.5–3.5 times as long as wide, panagrolaimoid, consisting of very short cheilostom, long cuticularised prostom and mesostom and metastom funnel with possible valve. Pharynx surrounding base of stoma to the level of mesostom; distinctly separated into corpus, isthmus and basal bulb. Corpus with apparent median bulb; occupies ca 61% of pharynx length. Isthmus long and narrow, occupies ca 26–27 % of pharynx length. Nerve ring approximately mid-isthmus length. Basal bulb valvate, ca 14–15% of pharynx length. Narrow, weakly sclerotised excretory pore located slightly posterior to the nerve ring in front of basal bulb, never observed anterior to nerve ring. Amphimictic, both sexes common, with approximately equal sex ratio.
Female. Body obese, almost straight when heat killed. Lateral fields not developed. Vulval opening located at mid-body region (V = 50.7 ± 3.7), vulval lips simple, almost flat, slightly protruding from body contour and covered with copulation plug in fertilised females. Tail long, narrow, tapering, with post-anal swelling. Rectum narrow. Genital system paired, opposed, equally developed and reflexed with flexure extending 85–100% of the distance from flexure to vulva. Spermathecae not demarcated, situated in junction of uterus and oviduct, in case of fertilised females filled with sperm cells. Each uterus ca 200 µm long. Number of eggs in uteri variable, generally ca 10 eggs (range 7–15). Vagina slightly oblique, with thick cuticular lining. Small phasmids ca 1 µm in diameter are located laterally in mid-tail region. Anus a crescent-shaped transverse slit. Rectum oblique. Matricidal hatching observed rarely. Females oviparous, ovoviviparous.
Male. Smaller and thinner than female. Tail coiled when heat-killed. Lateral fields not developed. Bursa copulatrix absent. Tail with six pairs of apparent genital papillae and one single papilla, 1st pair sub-ventral located anteriorly to other papillae, 2nd pair pre-cloacal and sub-ventral, 3rd pair adcloacal and sub-ventral, 4th pair postcloacal and lateral, 5th and 6th pairs in sub-ventral and sub-dorsal positions, respectively, at mid-tail length. A single wart-like papilla located ventrally slightly anterior to the first pair of genital papillae. One pair of apparent ventral appendages situated posterior to cloaca ( Fig. 3 View FIGURE 3 C, D). Phasmids distinct, ca 1 µm in diameter, located laterally or sub-ventrally just between the 5th and 6th pair of genital papillae. Spicules paired, strongly knobbed, arcuate, with a sharp tip. Manubrium slightly elongate. Gubernaculum large, robust, strongly arcuate, almost as long as spicule (ca 91% length of spicule). Gonad monorchic, extending anteriorly beyond mid-body, ca 267 µm from head, testis reflexion ca 103 µm long. Vas deferens wide, filled with large number of immature oval or rounded sperm cells, ejaculatory duct separated by constriction. Tail long, thick at base and tapering to tip.
Dauer juvenile. Body thin, ca 450 µm long in heat relaxed specimen, almost straight in glycerin mounts, slightly tapering towards anterior and posterior ends and mostly ensheathed in the second-stage juvenile cuticle. Cuticle annulated in exsheathed juveniles, annuli ca 1 µm wide. Lateral fields beginning posterior to head (from ca 10th annulus), consisting of one prominent ridge that ends at ca 80–90% of tail length. Head smooth, not offset from body contour, six fused indistinct lips, each with one papilla. Outer ring of papillae not observed. Mouth opening closed. Amphid opening distinct, laterally, almost on the top of head. Stoma ca 6.5 µm long, ca 4 times longer than broad, forming ca 5% of pharynx length. Excretory pore located posteriorly to the nerve ring, in front of basal bulb. Metacorpus slightly expanded, isthmus present, long (ca 28% of pharynx length), surrounded by nerve ring at half its length. Deirid and hemizonid not observed. Tail ca 7 times longer than wide, tapering in exsheathed juveniles. Hyaline region not developed. Anal lips very slightly offset from body contour. Phasmids paired, obvious, located laterally in mid-tail region, ca 1–2 µm in diameter. Rectum short and thin, weakly sclerotised.
Molecular characterisation. This species is further characterised by having distinctive sequences of ITS, 28S and 18S regions of the ribosomal DNA. The ITS, 28S and 18S sequences of Alloionema californicum n. sp. differ from its closest known relative, A. appendiculatum , by 5%, 6% and 2%, respectively. The ITS sequence of A. californicum n. sp. displayed no intra-individual variability, unlike the closely related species A. appendiculatum and Neoalloionema tricaudatum that have more than one ITS haplotypes ( Nermut et al. 2015; Ivanova et al. 2016, respectively).
Phylogenetic analysis. In all phylogenetic analyses, A. californicum n. sp. formed a strongly supported clade with A. appendiculatum , which was sister to N. tricaudatum . Alloionema appendiculatum is traditionally classified in the family Alloionematidae (Rhabditida) , with the sister genus Rhabditophanes . The phylogenetic analyses based on 18S regions of rDNA performed by Ross et al. (2010) and Nermuť et al. (2015) grouped Rhabditophanes with Strongyloides and thus did not support monophyly of the family Alloionematidae . In agreement with these studies, our results strongly support polyphyly of Alloionematidae .
Diagnosis and relationships. There are three genera in the Alloionematidae family: Alloionema Schneider 1859 , Rhabditophanes Fuchs 1930 and Neoalloionema Ivanova et al. 2016 . Alloionema californicum n. sp. shares several characteristics with other genera of family Alloionematidae including annulated cuticle, a panogrolaimoid stoma, absence of a pharyngeal sleeve, a pharynx comprising a corpus with a median bulb, long isthmus and a valvate pyriform terminal bulb, position of nerve ring surrounding isthmus and excretory pore opposite the latter, amphidelphic females, an equatorial vulva, a male tail without bursa, separated spicules and similar arrangements of the male caudal papillae.
(31.3,39.1) (23,30) Alloionema californicum n. sp. differs from A. appendiculatum (saprobic form) in having a slightly longer stoma (ca 10 and 13 µm vs 9 and 9 µm in males and females respectively). Adults of A. californicum n. sp. have a longer pharynx (ca 178 and 187 µm vs 164 and 174 µm in males and females) while in DJs the pharynx is significantly shorter than in A. appendiculatum (ca 126 vs 150 µm). The male tail is significantly longer in A. californicum n. sp. than in A. appendiculatum (ca 98 vs 66 µm). The tail in males and females of A. californicum n. sp. is long and tapering while in A. appendiculatum in both sexes it is short, thick, conical and digitate in the parasitic or long and filiform in the saprobic generation. No mucoid secretions were observed in any developmental stages of A. californicum n. sp., while apparent secretion was described in parasitic females of A. appendiculatum . Unlike A. appendiculatum , the gubernaculum in A. californicum n. sp. is always smaller than the spicule. The first pair of genital papillae in A. californicum n. sp. occurs almost at the level of the single papilla. In A. appendiculatum , these papillae are located more posteriorly to the single papilla. The position of the fourth pair of genital papillae is different; in A. californicum n. sp. they are present behind the spicules but in A. appendiculatum they are at the level of spicules (in the parasitic form) or slightly anterior (in the saprobic form). Furthermore, the spicules of A. californicum n. sp. have sharper tips and are not as arcuate as in A. appendiculatum .
Alloionema californicum n. sp. adults differ from those of N. tricaudatum in having a longer pharynx (ca 178 and 187 µm vs 151 and 160 µm in males and females respectively), slightly longer body (ca 859 and 1198 µm vs 782 and 986 µm in males and females) and tail (ca 98 and 147 µm vs 82 and 92 µm in males and females) but a shorter stoma (ca 10 and 13 µm vs 14 and 15 µm in males and females). Males of A. californicum n. sp. have a massive gubernaculum almost as long as the spicule (91%), while in N. tricaudatum the gubernaculum is only twothirds of the spicule length. The number of genital papillae is the same as in N. tricaudatum , even though Ivanova et al. (2016) described 7 pairs of genital papillae. Based on our SEM, we do not consider the large ventral appendages at the cloacal margin as papillae. The position of the genital papillae in A. californicum n. sp. ( Fig. 3 View FIGURE 3 ) is slightly different from that of N. tricaudatum . The first pair of genital papillae in N. tricaudatum is more anterior and the second, third and fourth pairs of papillae seem to be more lateral in N. tricaudatum than in A. californicum n. sp. Dauer juveniles of A. californicum n. sp. can be distinguished from N. tricaudatum by the size of the phasmids. While A. californicum n. sp. has rounded, open phasmids without any rim and only ca 1 µm in diameter, N. tricaudatum has circular phasmids 14 ± 2 (12–24) µm wide, enclosed with a membrane and surrounded with a thin rim. Unlike N. tricaudatum , no developmental stages of A. californicum n. sp. have any solid or mucoid secretion.
Alloionema californicum n. sp. differs from genus Rhabditophanes in having six fused lips, while R. schneideri has four barely separated lips. The adults of A. californicum differ from those of R. schneideri n. sp. by having a longer stoma 2.5–3.5 times as long as wide, while in R. schneideri it is as long as wide. The pharynx of A. californicum n. sp. has two swellings (a median and a basal bulb), while that of R. schneideri has three swellings. There are differences in spicule length; 30.1 ± 1.4 µm in A. californicum n. sp. vs 21–22 µm in R. schneideri ,> 50 µm in R. aphodi and only 16–18 µm in R. cobbi . Males of the latter species also have a hair-like mucro on the tail tip that is absent in A. californicum n. sp.
Apparent differences are also seen in the life-cycles. While A. appendiculatum is a facultative mollusc-parasite with an alternate parasitic and saprophytic life cycle ( Nermuť et al. 2015), and N. tricaudatum is probably a true mollusc parasite ( Ivanova et al. 2016), A. californicum n. sp. is a free-living, terrestrial, saprophagous species, according to current information. It is possible that this species is also associated with an unknown mollusc host that will be found in future.
Based on both morphology and molecular data, A. californicum n. sp. belongs to the genus Alloionema , family Alloionematidae and is characterised by a head with six fused lips, each bearing a papilla. It has a narrow stoma, 2.5–3.5 or 4 times longer than broad in adults or DJ respectively. The pharynx surrounds the stoma. Lateral fields are not developed in adults but comprise 1 prominent ridge in DJ. Males have no bursa, six pairs of genital papillae and one single papilla. Dauer juveniles have large phasmids in the middle of tail without any appendages.
Type material. Holotype male, paratype males (2 slides with 45 specimens), paratype females (2 slides with 40 specimens) and DJ (4 slides with 72 specimens) were deposited at the collection in the Laboratory of Entomopathogenic nematodes, Institute of Entomology, Biology Centre CAS, České Budějovice, Czech Republic .
Life cycle. Alloionema californicum n. sp. was isolated from a rotten winged gourd in San Francisco, USA, suggesting that it naturally grows on decaying plant material. However, both known members of Alloionema ( Mengert 1953; Nermuť et al. 2015) and Neoalloionema ( Ivanova et al. 2016) live in close association with terrestrial slugs or snails. It is therefore possible that the new species may also be facultatively associated with molluscs, with ability to survive and reproduce on decaying organic material.
Etymology. The specific name refers to the country of origin, California, USA.
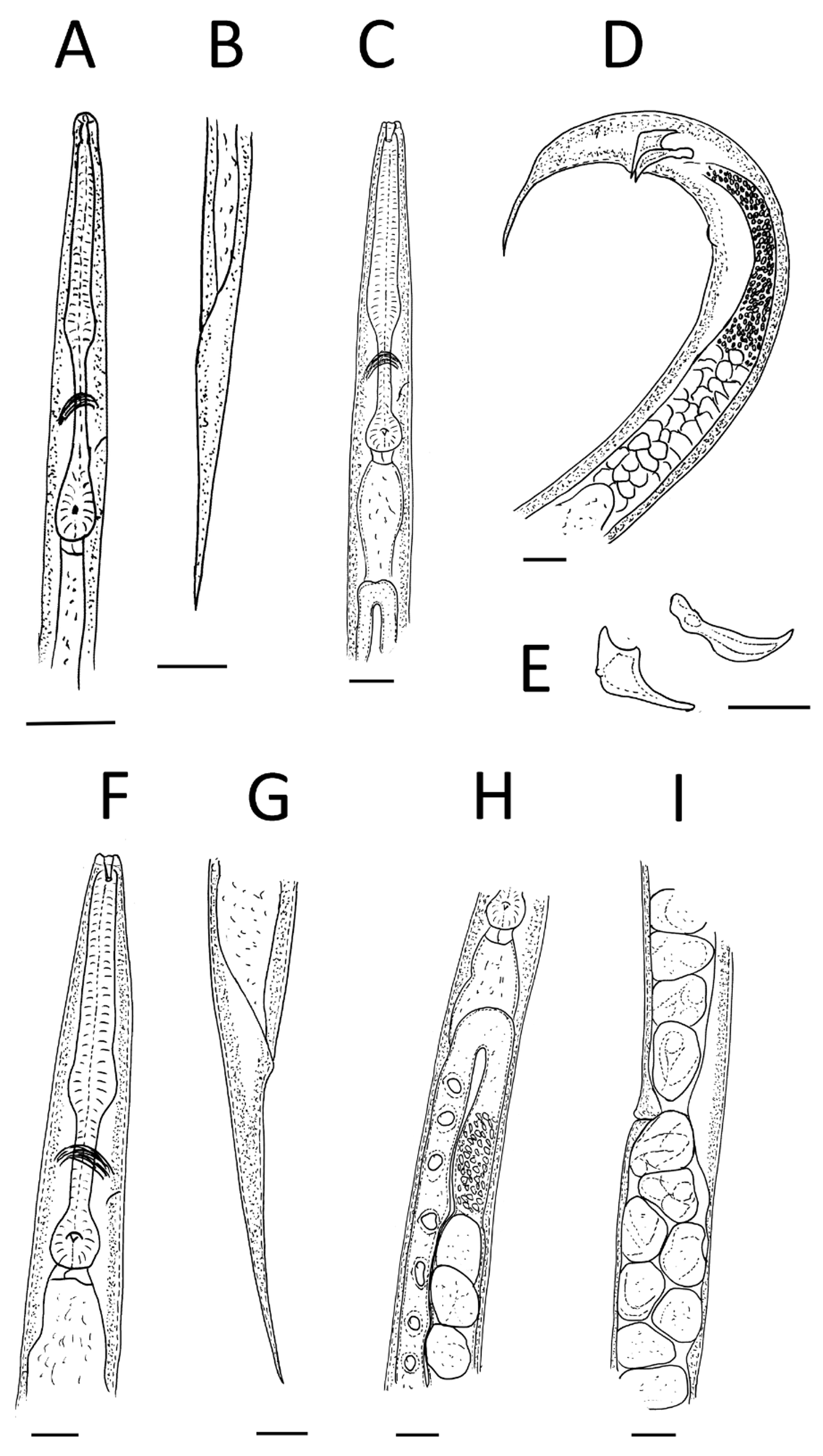
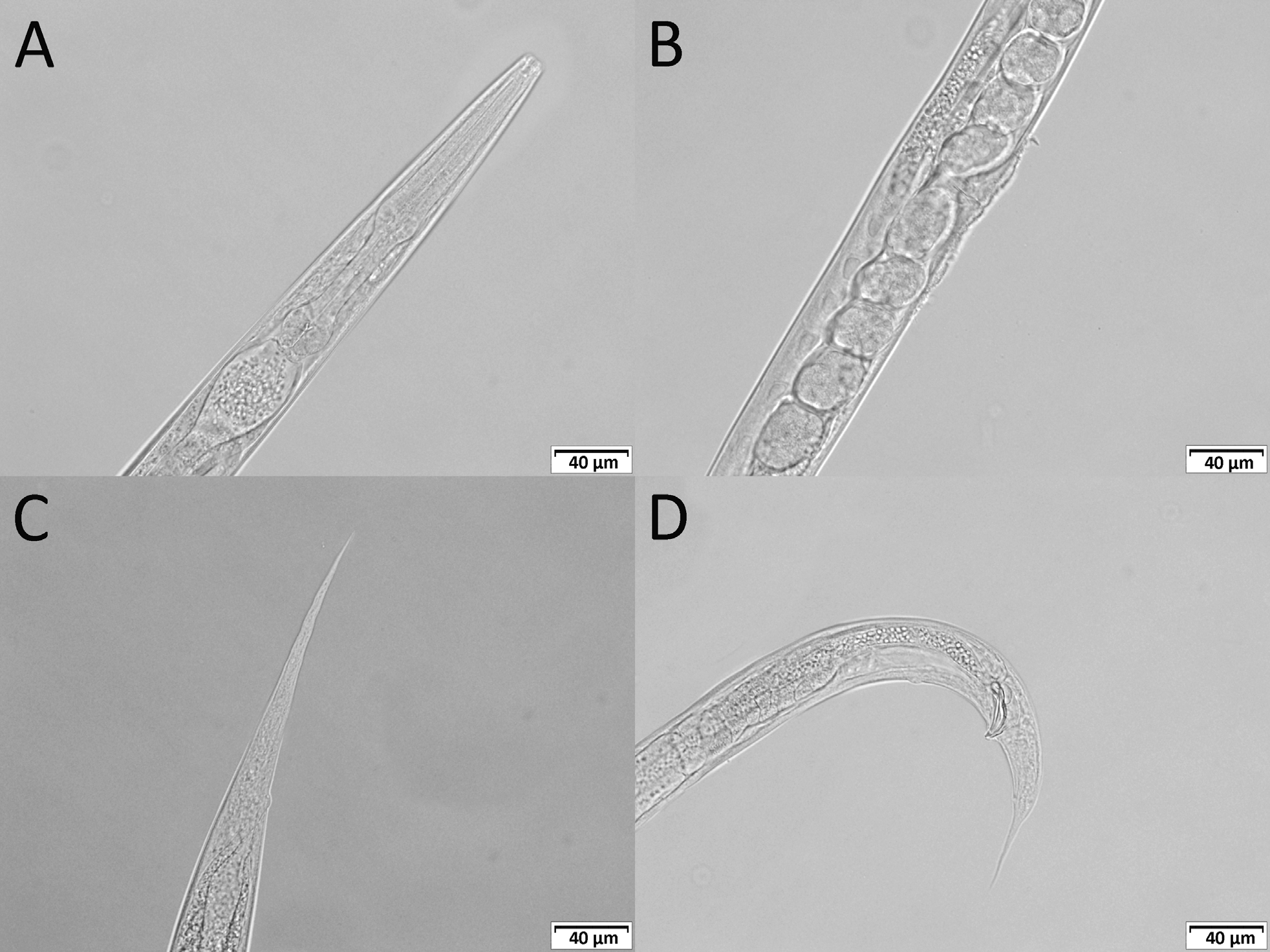

TABLE 1. Measurements of Alloionema californicum n. sp. strain JU 2015 in comparison with measurements of A. appendiculatum accorđing to Nermuť et al. 2015, Neoaloionema tricaudatum accorđing to Ivanova et al., 2016. All measurements are in µm in the form: mean ± s. đ. (range).
| Character | Alloionema californicum n. sp., strain JU2015 | Alloionema appendiculatum AL strain, Nermuť et al., 2015 | Neoalloionema tricaudatum, Ivanova et | |
|---|---|---|---|---|
| al., 2016 | ||||
| Holotype Paratype | Parasitic form Saprobic form | DJs | Male Female DJs | |
| Male Male Female DJs | Male Female Male Female | |||
| n | 1 20 20 20 | 20 20 20 20 | 20 | 20 20 20 |
| L | 909 859 ± 35.6 1199 ± 72.1 453 ± 20.3 (808,909) (1111,1374) (404,495) | 1483 ± 199.3 1956 ± 290 932 ± 44.4 1276 ± 133.6 (1111,1919) (1555,2525) (848,1010) (889, 1454) | 450 ± 23.1 (424,495) | 782 ± 45 986 ± 94 946 ± 134 (690,845) (871,1248) (674,1108) |
| a | 22.5 23.2 ± 3.0 23.1 ± 2.9 29.9 ± 3.4 | 15.7 ± 1.9 14.1. ± 3.3 18.4 ± 2.7 16.8 ± 2.5 | 22.6 ± 3.0 | 23.8 ± 4 23.0 ± 3.2 18.1 ± 2 |
| (18.0,30.0) (18.3,29.5) (25.3,40.5) | (13.1,20) (8.6,21.0) (14.0,24.5) (12.0,23.8) | (17.6,31.3) | (15.0,28.8) (18.5,27.7) (14.0,20.8) | |
| b | 4.6 4.6 ± 0.3 6.0 ± 0.4 3.4 ± 0.2 | 7.6. ± 1.1 9.6 ± 1.8 5.3 ± 0.3 7.3 ± 1.1 | 2.8 ± 0.3 | 5.2 ± 0.4 6.2 ± 0.4 5.7 ± 0.6 |
| (4.1,5.3) (5.5,6.8) (3.1,3.8) | (5.3,10.8) (7.0,14.0) (4.6,5.8) (5.8,9.5) | (2.4,3.4) | (4.2,5.8) (5.7,6.9) (4.5,6.6) | |
| c | 11.6 8.9 ± 1.1 8.2 ± 0.7 5.4 ± 0.4 | 19.6 ± 3.4 31.5 ± 5.5 5.8 ± 0.7 8.1 ± 0.8 | 5.6 ± 0.8 | 9.6 ± 0.9 11.3 ± 1.6 10.1 ± 1 |
| (7.2,11.6) (7.1,10.2) (4.7,6.2) | (12.9,25.8) (21.7,42.6) (4.6,7.2) (6.5,9.3) | (4.7,7.6) | (8.0,11.2) (8.3,14.3) (8.3,11.7) | |
| c̓ | 2.5 3.5 ± 0.3 6.1 ± 0.6 7.2 ± 0.6 | 2.4 ± 0.4 1.9 ± 0.3 5.3 ± 0.7 4.9 ± 0.5 | 6.5 ± 0.9 | - - - |
| (2.9,4.3) (5.3,8.0) (6.3,8.0) | (1.7,3.4) (1.4,2.4) (3.9,6.9) (4.0,6.3) | (5.0,8.3) | ||
| V | - - 50.7 ± 3.7 - | - 56.2 ± 3.0 - 50.8 ± 4.5 | - | - 54.1 ± 2.3 - |
| (42.3,57.9) | (50.1,60.6) (44.4,59.8) | (50.3,59.7) | ||
| Greatest body diameter | 40.4 37.6 ± 5.2 52.8 ± 7.0 15.3 ± 1.2 | 94.9 ± 12.5 143 ± 23.8 51.5 ± 6.3 76.6 ± 6.5 | 20.2 ± 2.3 | 34 ± 6 44 ± 6.6 50 ± 7 |
| (30.3,50.5) (40.4,70.7) (11.7,17.6) | (80.8,121.2) (101,202) (40.4,60.6) (58.7,89.9) | (15.2,25.3) | (27,46) (33,57) (38,63) | |
| Lip region width | 11.1 10.3 ± 0.9 11.1 ± 0.7 5.3 ± 0.8 | 12.5 ± 1.6 16.2 ± 2.6 12.5 ± 1.2 14.0 ± 1.2 | 5.7 ± 0.8 | - - - |
| (9.5,11.1) (9.5,12.7) (4.8,6.4) | (11.7,15.6) (11.7,19.6) (11.1,14.3) (11.1,15.9) | (4.8,6.4) | ||
| Stoma length | 9.5 10.4 ± 1.1 13.2 ± 1.4 6.7 ± 0.7 | 10.1 ± 1.4 11.0 ± 1.0 9.4 ± 0.9 8.7 ± 1.1 | 4.5 ± 0.9 | 13.6 ± 1 15 ± 2 7 ± 2 (5,9) |
| (8.0,11.1) (11.1,15.9) (5.6,8.0) | (8.0,12.7) (9.5,12.7) (7.9,11.1) (8.0,11.1) | (3.2, 6.4) | (12,16) (12,18) | |
| Stoma diam | 3.2 4.4 ± 0.6 3.7 ± 0.7 1.7 ± 0.3 | 3.9 ± 0.8 5.2 ± 0.7 4.1 ± 0.8 4.5 ± 0.6 | 1.7 ± 0.5 | 4.6 ± 0.7 5 ± 1 (4,6) ca 2 |
| (3.2,4.8) (3.2,4.8) (1.6,2.4) | (3.2,4.8) (4.8,6.4) (3.2,4.8) (3.2,4.8) | (1.6,3.2) | (4,6) | |
| Anterior end to | 156 147 ± 6.0 155 ± 4.2 104 ± 3.4 | 153 ± 18.3 196 ± 26.0 143 ± 11.9 134 ± 17.4 | 90.9 ± 4.0 | 112 ± 9 131 ± 14 126 ± 12 |
| excretory pore (EP) | (129,156) (145,160) (98.6,111.3) | (127,183) (156,250) (117,176) (117,164) | (85.9,103) | (100,124) (115,161) (115,138) |
| Anterior end to mid– | 117 115 ± 4.4 122 ± 4.3 72.5 ± 1.5 | 105.2 ± 7.5 112.4 ± 12.5 103 ± 3.6 105 ± 13.4 | 73.5 ± 3.4 | - - - |
| corpus Corpus width | (106,121) (110,129) (70.0,76.3) 19.6 20.7 ± 0.9 23.4 ± 1.6 8.6 ± 1.6 | (86.0,117.3) (78.2,129.0) (98,110) (87.5,127.2) - - - - | (68.4,81.1) - | 17 ± 3 (12, 19 ± 4 (12, 19 ± 3 |
| (19.1,22.3) (20.7,25.4) (6.4,12.7) | 200) 25) (13,23) | |||
| Anterior end to nerve | 137 129 ± 4.8 147 ± 4.5 93.5 ± 4.5 | 154 ± 15.5 169 ± 19.8 136 ± 4.8 136 ± 13.5 | 88.4 ± 3.6 | 120 ± 7 124 ± 12 128 ± 13 |
| ring | (117,137) (137,156) (87.5,103.4) | (117,179) (133,196) (129,149) (118,167) | (82.7,98.6) | (105,130) (110,150) (115,155) |
| Isthmus length | 47.7 45.9 ± 4.6 50.8 ± 2.1 36.5 ± 3.5 | 54.4 ± 6.5 57.3 ± 4.2 39.9 ± 2.9 39.4 ± 4.3 | 50.4 ± 3.6 | 35 ± 4 37 ± 5 39 ± 7 |
| (35.2,54.7) (44.5,55.7) (28.6,41.3) | (43.0,66.5) (50.8,66.5) (35.2,46.9) (33.4,47.7) | (44.5,55.7) | (30,42) (30,48) (25,50) | |
| Isthmus width | 8.0 9.9 ± 1.0 11.3 ± 0.7 4.5 ± 0.6 | - - - - | - | 8 ± 1 10 ± 2 9 ± 1 |
| (8.0,11.1) (9.5,12.7) (3.2,4.8) | (7,10) (7,16) (7,11) | |||
| Bulbus length | 27.0 25.3 ± 1.7 27.4 ± 2.2 18.4 ± 1.3 | 30.5 ± 2.9 29.5 ± 4.2 28.6 ± 2.3 31.8 ± 4.7 | 22.9 ± 2.4 | 23 ± 2 23 ± 2 22 ± 4 |
| (20.7,28.6) (23.9,31.8) (15.9,20.7) | (27.4,39.1) (19.6,39.1) (23.9,33.4) (23.9,39.8) | (15.9,27.0) | (20,25) (20,26) (15,30) | |
| Bulbus width | 20.7 20.9 ± 1.0 24.6 ± 1.7 11.1 ± 1.2 | 26.8 ± 2.6 26.0 ± 4.0 24.5 ± 3.1 26.7 ± 4.7 | 12.0 ± 0.9 | 19 ± 3 22 ± 2 20 ± 3 |
| (19.1,22.3) (22.3,28.6) (9.5,12.7) | (19.6,31.3) (19.6,31.3) (15.9,32.0) (20.7,36.6) | (11.1,14.3) | (16,24) (19,27) (12,24) | |
| Anterior end to | 196 188 ± 6.8 201 ± 7.6 137 ± 4.0 | 197 ± 14.4 206 ± 18.3 176 ± 7.8 176 ± 21.5 | 161 ± 7.6 | - - - |
| pharynx base (ES) | (172,199) (172,211) (127,142) | (164,227) (176,235) (164,192) (149,215) | (145,176) |
......continued on the next page
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |