
Akodon kadiweu, Brandão & Percequillo & D’Elía & Paresque & Carmignotto, 2021
|
publication ID |
https://doi.org/ 10.1093/jmammal/gyaa126 |
|
publication LSID |
lsid:zoobank.org:pub:8E54E468-53E2-481B-93FF-EBB9168BFB05 |
|
persistent identifier |
https://treatment.plazi.org/id/AC08B99F-DCA0-4D6A-A3E1-3FAEC2010CF2 |
|
taxon LSID |
lsid:zoobank.org:act:AC08B99F-DCA0-4D6A-A3E1-3FAEC2010CF2 |
|
treatment provided by |
Felipe |
|
scientific name |
Akodon kadiweu |
| status |
sp. nov. |
Akodon kadiweu , new species
( Figs. 5–9 View Fig View Fig View Fig View Fig View Fig )
Holotype.— MZUSP 35766 View Materials , adult male (age-class 3), preserved as skin, skull, skeleton, and tissue in alcohol, collected by A. P. Carmignotto and colleagues on 19 April 2002 (original field number APC 985 ). Karyotype 2n = 40, FN = 40. Measurements listed in Table 2.
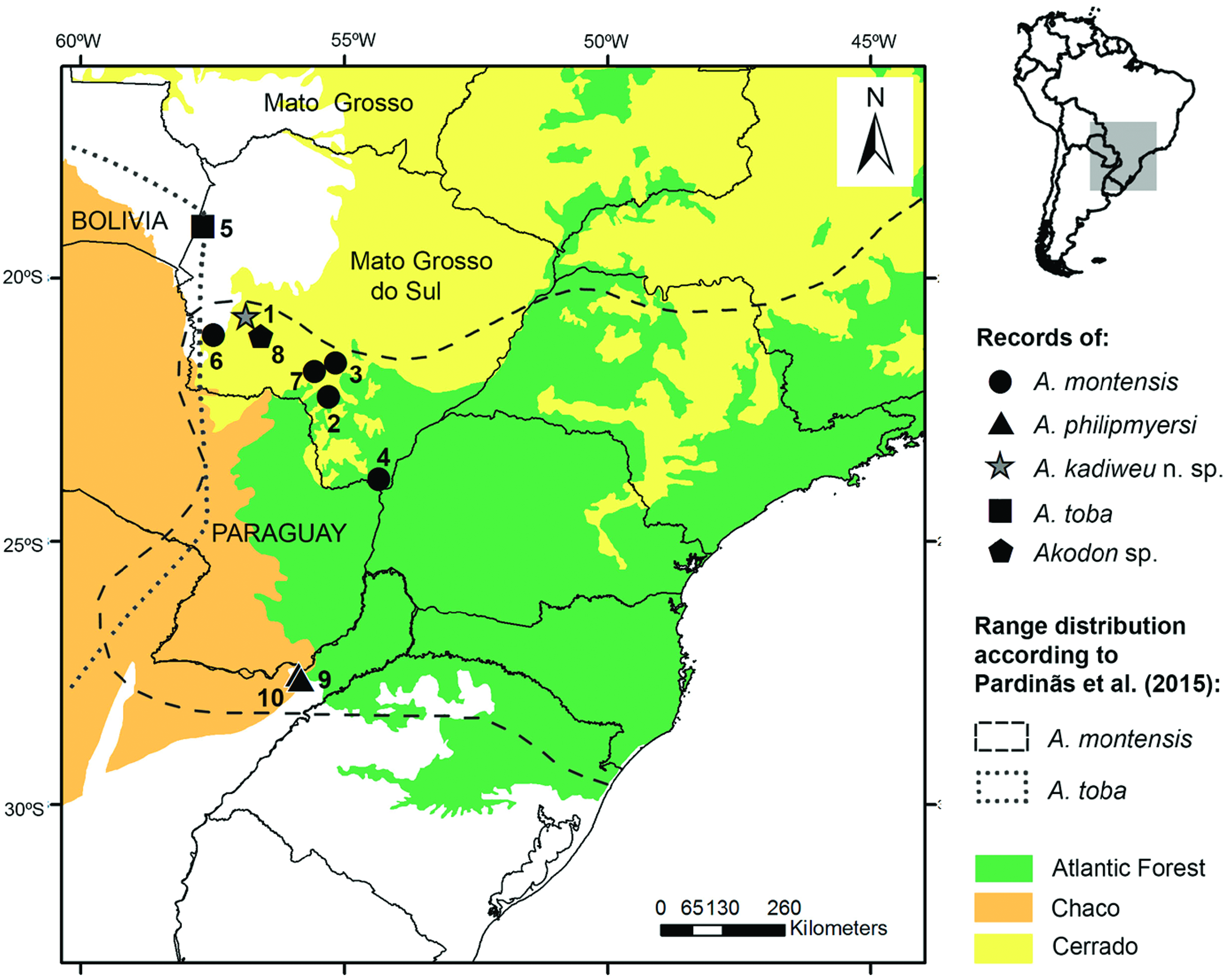
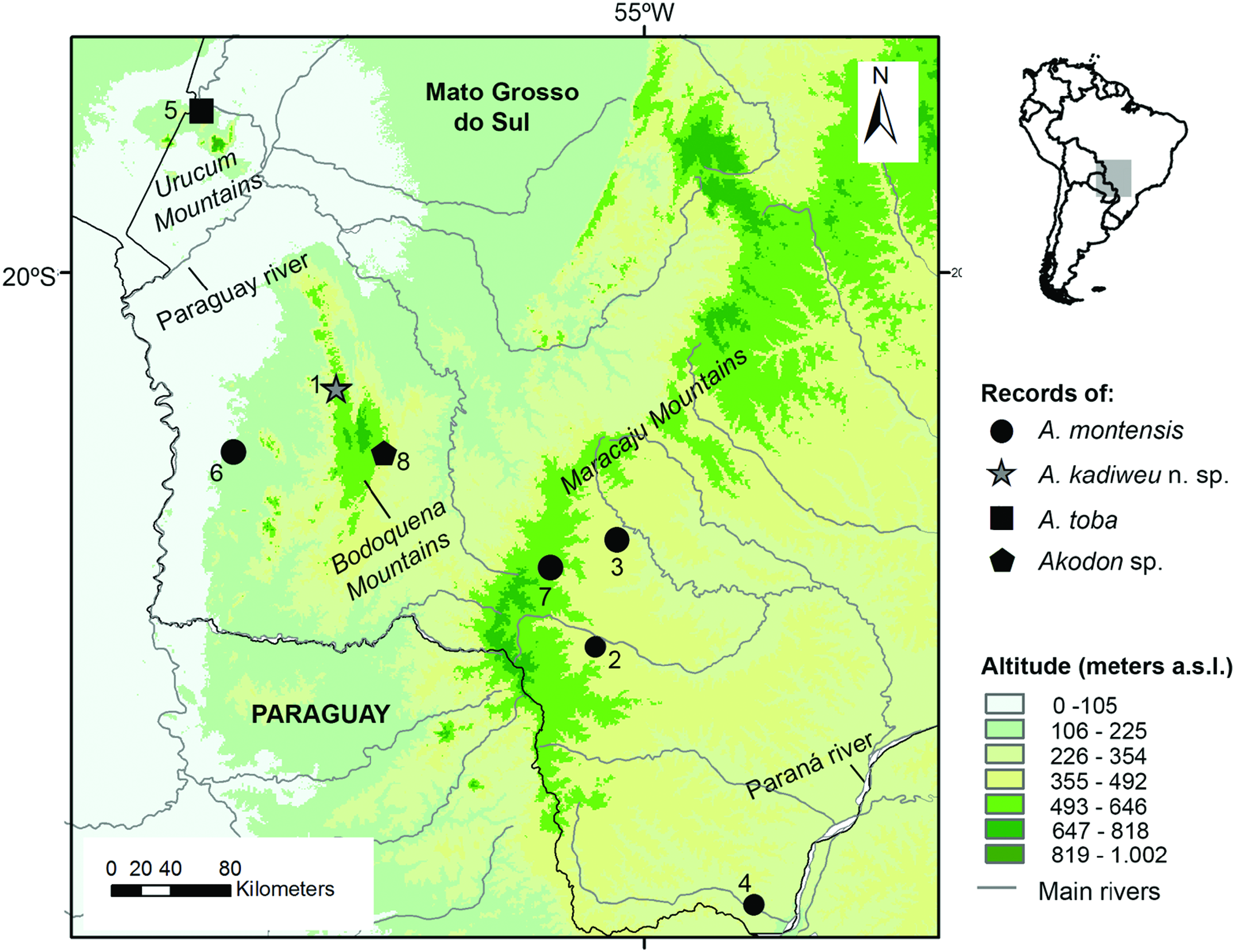
Type locality.— Brazil: Mato Grosso do Sul state; Bodoquena municipality, Serra da Bodoquena, Fazenda Califórnia , (20°42′S; 56°51′W; 520 m a.s.l.). This locality currently lies within the limits of the PNSB, a federal conservation unit within the Cerrado region , in southwestern GoogleMaps Mato Grosso do Sul state ( Fig. 1 View Fig ).
Paratypes.— Three additional specimens collected by the same expedition at the type locality during April 2002, including two males ( MZUSP 35765 View Materials , 35767 View Materials ), and a female ( MZUSP 35994 View Materials ). Sequences of 801 bp of the Cytb gene of these specimens have been deposited in GenBank with accession numbers MT273066 View Materials , MT273067 View Materials , and MT273068 View Materials , respectively. The paratype MZUSP 35765 View Materials has a karyotype of 2n = 40, FN = 40, similar to the holotype. Measurements listed in Table 2 .
Diagnosis.— This species is diagnosed by the following exclusive combination of traits: pelage dark-furred grayish-brown, medium-sized (HBL mean = 88.50 mm; CIL mean = 25.19 mm; MTRL mean = 4.42 mm; see Table 2); tail nearly as long as head and body length (TL mean = 82 mm; see Table 2); skull slender (RW/RL 48% mean), with moderately long rostrum (10.66 ± 0.20; 10.43–10.84); shallow zygomatic notch and narrow zygomatic plate (2.28 ± 0.28, 2.00–2.56); wide interorbital region and braincase (respectively: 5.09 ± 0.78, 5.04–5.18; and 12.24 ± 0.09, 12.18–13.25); incisive foramina reaching the protoflexus or anterior border of protocone of M1; palatal bridge with mesopterygoid fossa with anterior margin even with the posterior M3 level; anterior margin of the mesopterygoid fossa with a distinct medial posterior projection; conspicuous sphenopalatine vacuities; parapterygoid fossae wider than the mesopterygoid fossa; first lower molar with developed mesolophid, ectolophid, and ectostylid; chromosomal formula, 2n = 40 and FN = 40.
Distribution.— Known exclusively from the type locality, Fazenda California (in the limits of the PNSB), which is located in the northern portion of the Serra da Bodoquena, in the municipality of Bodoquena, Mato Grosso do Sul state, Brazil ( Fig. 10 View Fig ). Villages and farmlands characterize the surroundings of this conservation unit, with landscape altered by human activity, mainly deforestation for cattle ranching, a widespread activity in the area ( Carmignotto 2005; also see Oliveira et al. 2009).
Etymology.— Named after the indigenous people that live in the region where the new species is found, known as Kadiwéu ( Pechincha 1998; Silva 2018). Indigenous protected areas, such as the Reserva Indígena Kadiwéu nearby Serra da Bodoquena, have long been important for native people, as well as native vegetation and fauna preservation in South America ( Ricketts et al. 2010), for which reason we stand for their continuous maintenance and increase in Brazil.
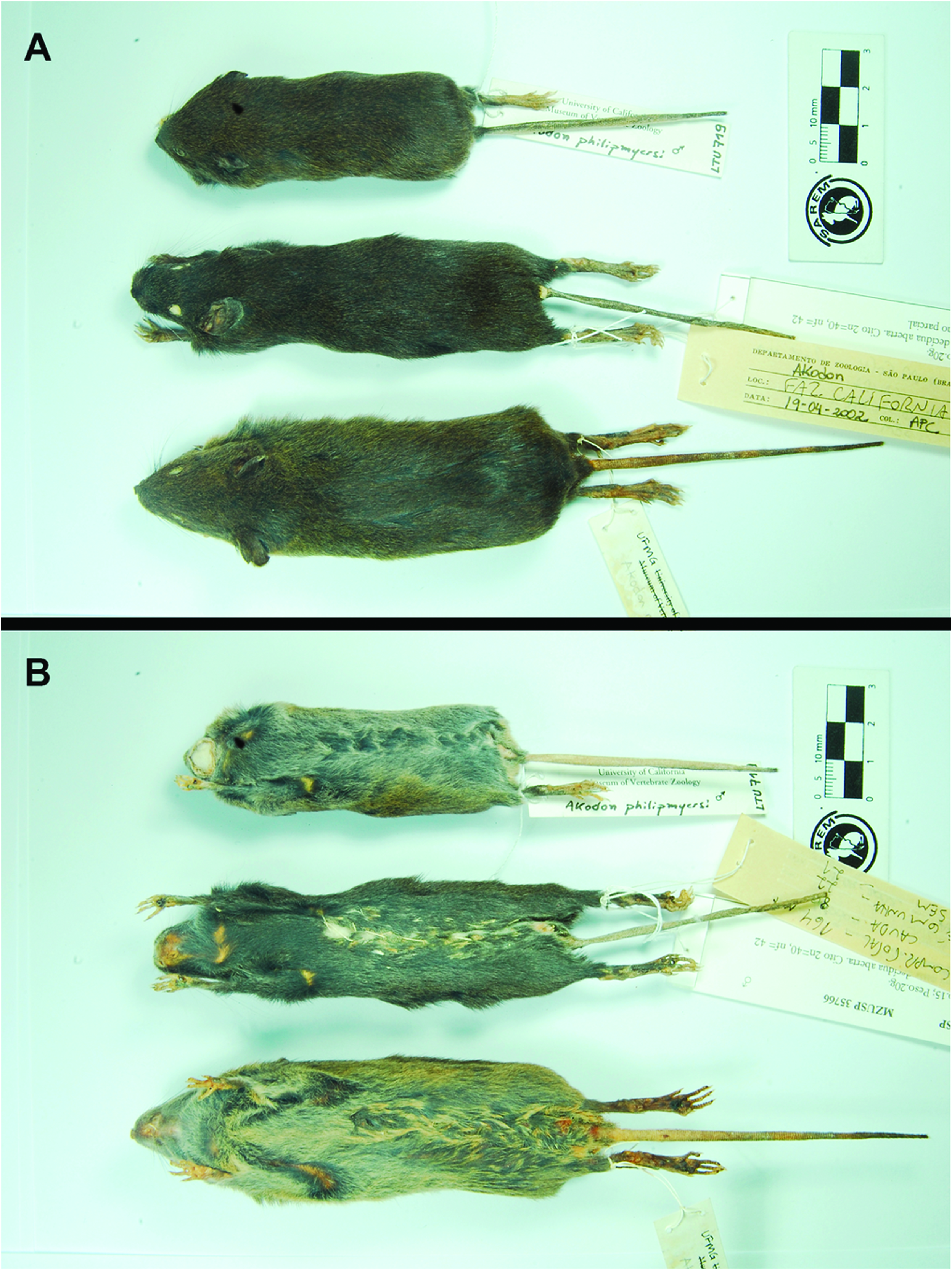
Morphological description.— Medium-sized Akodon (based on the range of species from eastern South America, with mean size ranging from 63 to 131 mm — Gonçalves et al. 2007), with moderate head and body length (HBL) and body mass (Table 2); tail length nearly as long as the HBL (87–97% of HBL; Supplementary Data SD1); pes moderately narrow and long (25–26% of HBL; Supplementary Data SD2); pinnae prominent (17–19% of HBL) and covered by delicate and short brownish hairs (Supplementary Data SD3). Pelage dark gray-based; dorsum covered by soft and moderately long hairs (mean length: 10 mm), dark grayish-brown slightly grizzled with black guard hairs (from crown to rump); ventral region homogeneously dark gray; gradual transition between grayish-brown dorsal coloration to grayish ventral coloration ( Fig. 5 View Fig ). Eye-ring present only in one of the four specimens (MZUSP 35994). Mystacial vibrissae moderately long, reaching but not surpassing pinnae when laid back, and blackish and/or silvery colored (Supplementary Data SD3). Tail bicolored, covered with short and sparse blackish hairs dorsally and whitish hairs ventrally, scales small but visible to the naked eye, regular, and with 1–3 hairs per scale. Dorsal surface of manus and pes buffy-brown, sparsely covered by a mixture of silvery and blackish hairs, with moderately long claws (1–2 mm), all covered by silvery ungual tufts as long (or slightly longer) as claws; ventral surface naked and unpigmented. Manus short and with two carpal and three plantar pads. Pes relatively long (2.25 ± 0.5, 22.0–23.0) and narrow ( Fig. 5 View Fig ), with outer digits (I and V) much shorter than the central ones (II, III, and IV, all similar-sized); ventral surface with poorly distinct squamae (scale-like tubercles) sparsely distributed, and with moderately large and fleshy interdigital (1–4) and tarsal pads (thenar and hypothenar; Supplementary Data SD2); all pads are well-developed, but with different sizes: pads 2 and 3 are the largest ones; followed by pad 4 and thenar, which are subequal in size; and pad 1 and hypothenar, the smallest ones.
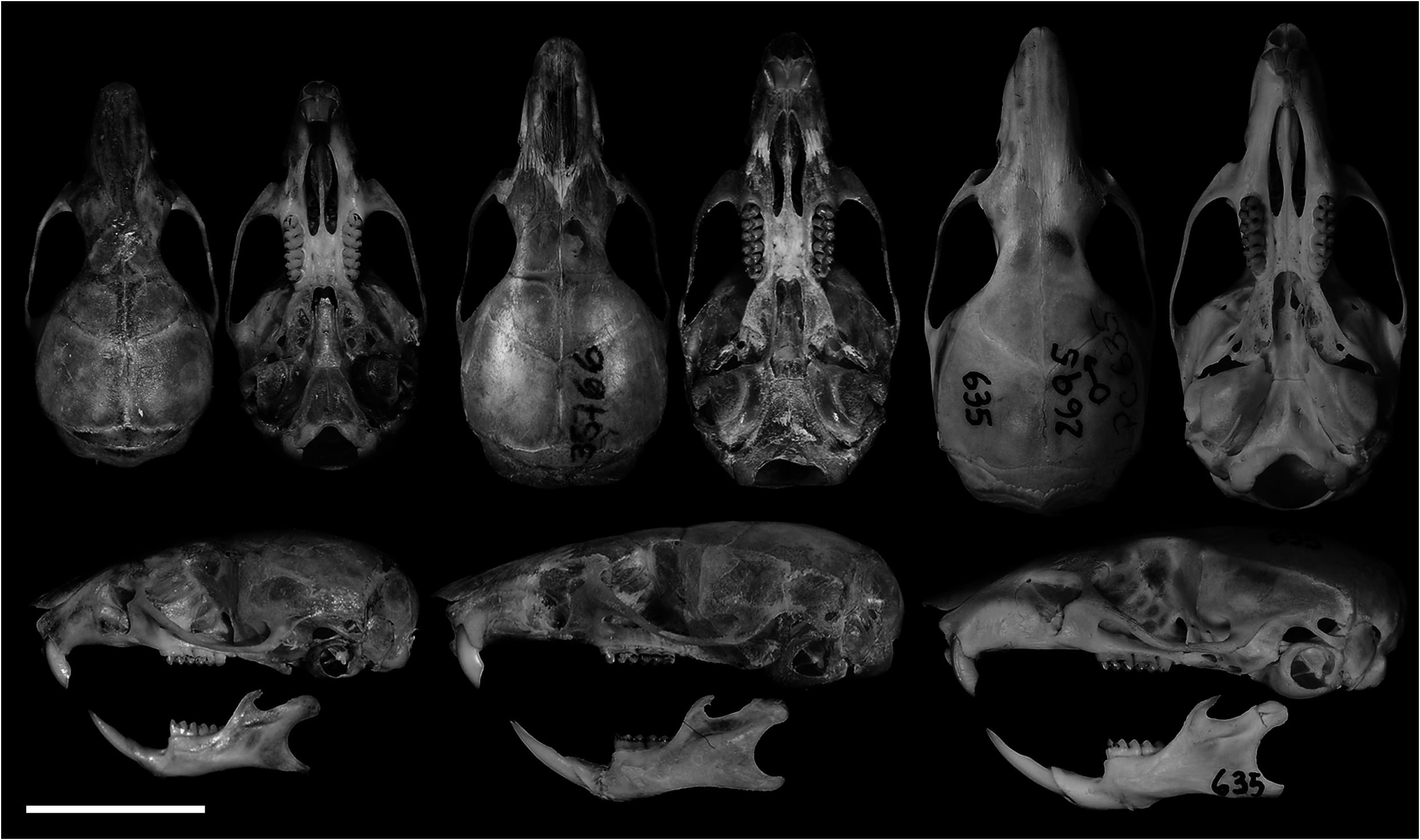
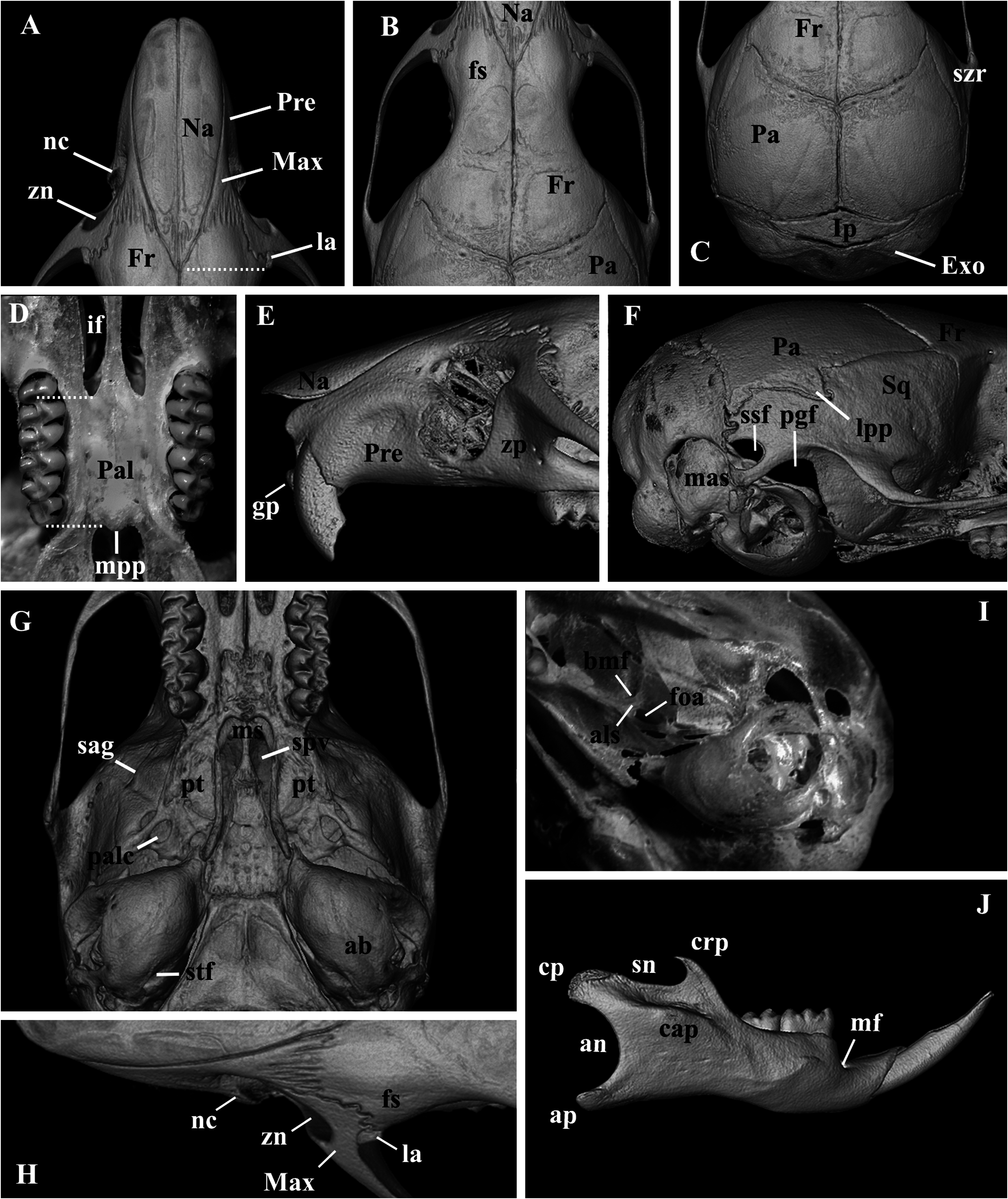
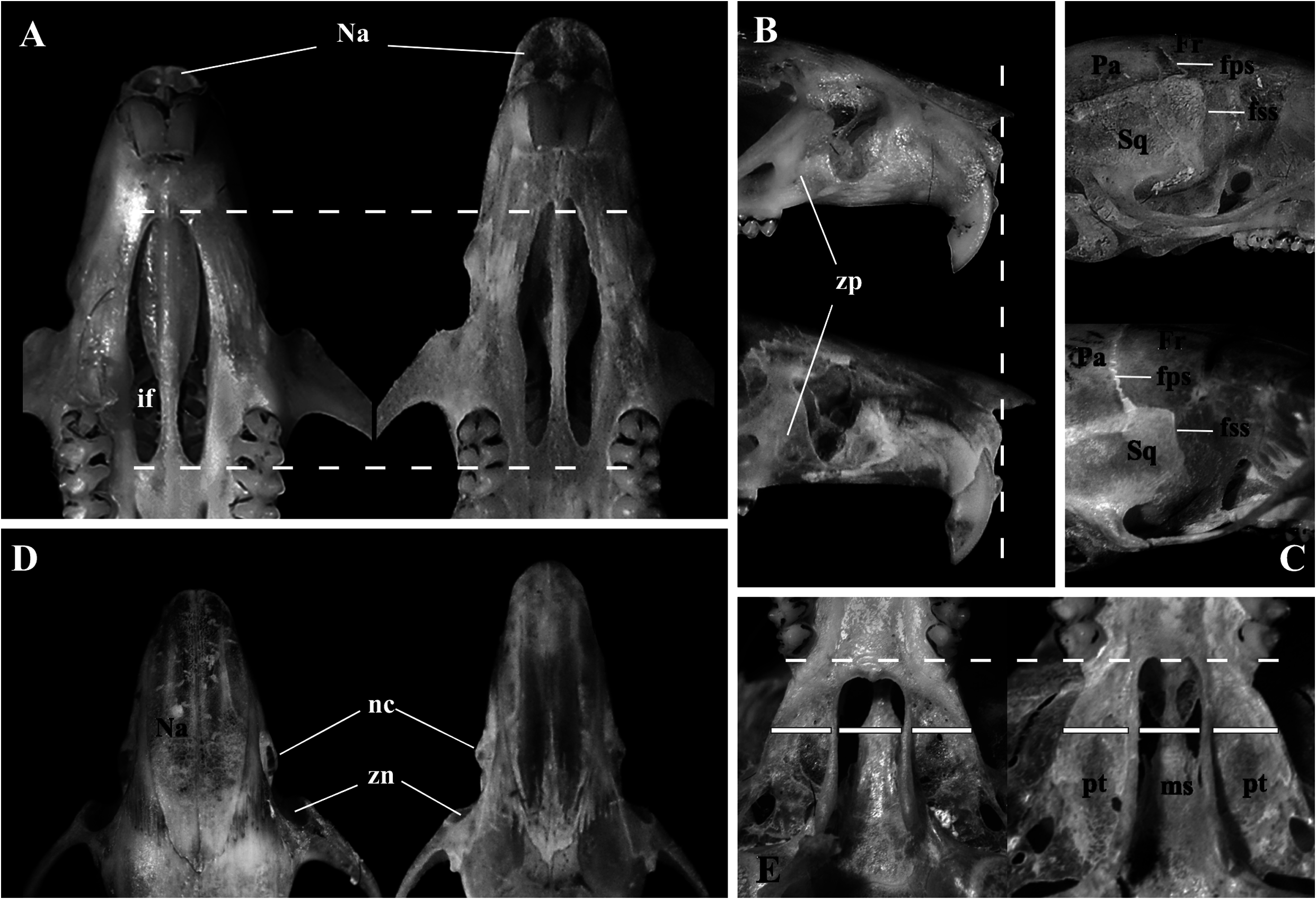
Skull (in dorsal view) characterized by a moderately long rostrum (10.66 ± 0.20; 10.43–10.84; Figs. 6 View Fig and 7A View Fig ); rostrum with poorly developed capsular projection of nasolacrimal foramen; rostrum flanked by shallow and narrow zygomatic notches ( Fig. 7H View Fig ) due to a narrow zygomatic plate (2.28 ± 0.28, 2.00–2.56); nasals moderately wide, not covering all the rostrum area in dorsal view ( Fig. 7A View Fig ), and long (10.42 ± 0.33, 10.04–10.68), extending anteriorly beyond the premaxillae ( Figs. 7E View Fig and 9B View Fig ) and acuminating at the posterior end, near the lacrimal level ( Fig. 7A View Fig ); gnathic process slightly projecting anteriorly the anterior incisors surface ( Fig. 7E View Fig ). Nasofrontal ( Fig. 7B View Fig ) and frontoparietal ( Fig. 7C View Fig ) sutures V-shaped. Interorbital region hourglass-shaped, wide (5.09 ± 0.78, 5.04–5.18), and with rounded supraorbital margins ( Fig. 7B View Fig ); frontal sinuses moderately inflated ( Figs. 7B and 7H View Fig ). Lacrimals moderately developed, equally in contact with maxillary and frontal ( Fig. 7H View Fig ). Braincase wide (12.24 ± 0.09, 12.18–13.25) and rounded, with no temporal beads or ridges; lambdoidal and nuchal crests absent or weakly developed ( Figs. 7C and 7F View Fig ); interparietal relatively narrow relative to its length, and with moderately long sutures between the parietal and exoccipital ( Fig. 7C View Fig ). Dorsal profile (in lateral view) slightly convex ( Fig. 6 View Fig ); premaxillae and nasals slightly projected anteriorly beyond the incisors; zygomatic plate slopes gently backward from bottom to top (slightly concave), with upper free border distinctly reduced and zygomatic spine absent ( Fig. 7E View Fig ); small masseteric scar present in front of the root of the zygomatic plate. Parietal slightly expanded onto lateral surface of braincase (posteriorly to the end of the squamosal zygomatic root; Fig. 7F View Fig ); frontosquamosal suture anterior to frontoparietal suture ( Figs. 7F View Fig and 9C View Fig ), which allows further contact between the squamosal and frontal. Zygomatic arches delicate (see comparisons to other akodontines in Missagia and Perini [2018]), formed by jugal, zygomatic process of maxillary, and zygomatic process of squamosal; zygomatic arches wider at the squamosal zygomatic roots and only slightly wider than braincase ( Fig. 7C View Fig ). Stapedial foramen, sphenofrontal foramen, and squamosal-alisphenoid groove present ( Fig. 7G View Fig ), configuring the primitive carotid circulatory pattern 1 (sensu Voss 1988). Alisphenoid strut consistently present in all specimens configuring discrete oval accessory and buccinator-masticatory foramina ( Fig. 7I View Fig ). Hamular process of squamosal wide, resulting in a moderately developed postglenoid foramen and subsquamosal fenestra, being the former slightly larger ( Fig. 7F View Fig ). Tegmen tympani overlapped to a small posterior suspensory process of squamosal; mastoid with a conspicuous fenestra ( Fig. 7F View Fig ). Incisive foramina wider anteriorly, near the premaxillae/maxillary suture, and long (6.30 ± 0.20, 6.18–6.54); averaging about 90% of length of diastema, with posterior margins extending to protoflexus ( Figs. 6 View Fig and 7D View Fig ) or anterior portion of protocone of M1 ( Fig. 9A View Fig ). Palate short and wide (sensu Hershkovitz 1962), with anterior margin of mesopterygoid fossa aligned to the posterior margin of M3; palate smooth, without pronounced traits ( Fig. 7D View Fig ), with shallow posterolateral pits in the oldest individual (age-class 5). Mesopterygoid fossa narrower than the parapterygoid fossae ( Fig. 7G View Fig ), with anterior margin lire-shaped, always with a prominent median palatine process ( Fig. 7D View Fig ); moderately wide sphenopalatine vacuities, perforating most of the bony roof of mesopterygoid fossa ( Fig. 7G View Fig ). Hamular process of pterygoid projected ventrally and slightly divergent at posterior end; parapterygoid fossae moderately excavated and wider than mesopterygoid fossa ( Fig. 7G View Fig ). Basioccipital poorly excavated; auditory bullae flask-shaped, with short but wide eustachian tube ( Fig. 7G View Fig ). Foramen magnum oval in shape; occipital condyles not projected posteriorly.
Mandible relatively short (14.67 ± 1.15, 14.0–16.0) and delicate, with anterior point of diastema located slightly below alveolar plane ( Fig. 7J View Fig ). Condyloid process moderately developed; coronoid process falciform, subequal in height to the condyloid process; angular process short, reaching the same level of condyloid process posteriorly ( Fig. 7J View Fig ). Sigmoid and angular notches moderately deep. Masseteric crest poorly developed, but superior and inferior ridges converge anteriorly as open chevron that ends below the middle or procingulum of ml level. Capsular process of lower incisors poorly developed, lying at base of coronoid process; mental foramen visible in lateral view ( Fig. 7J View Fig ).
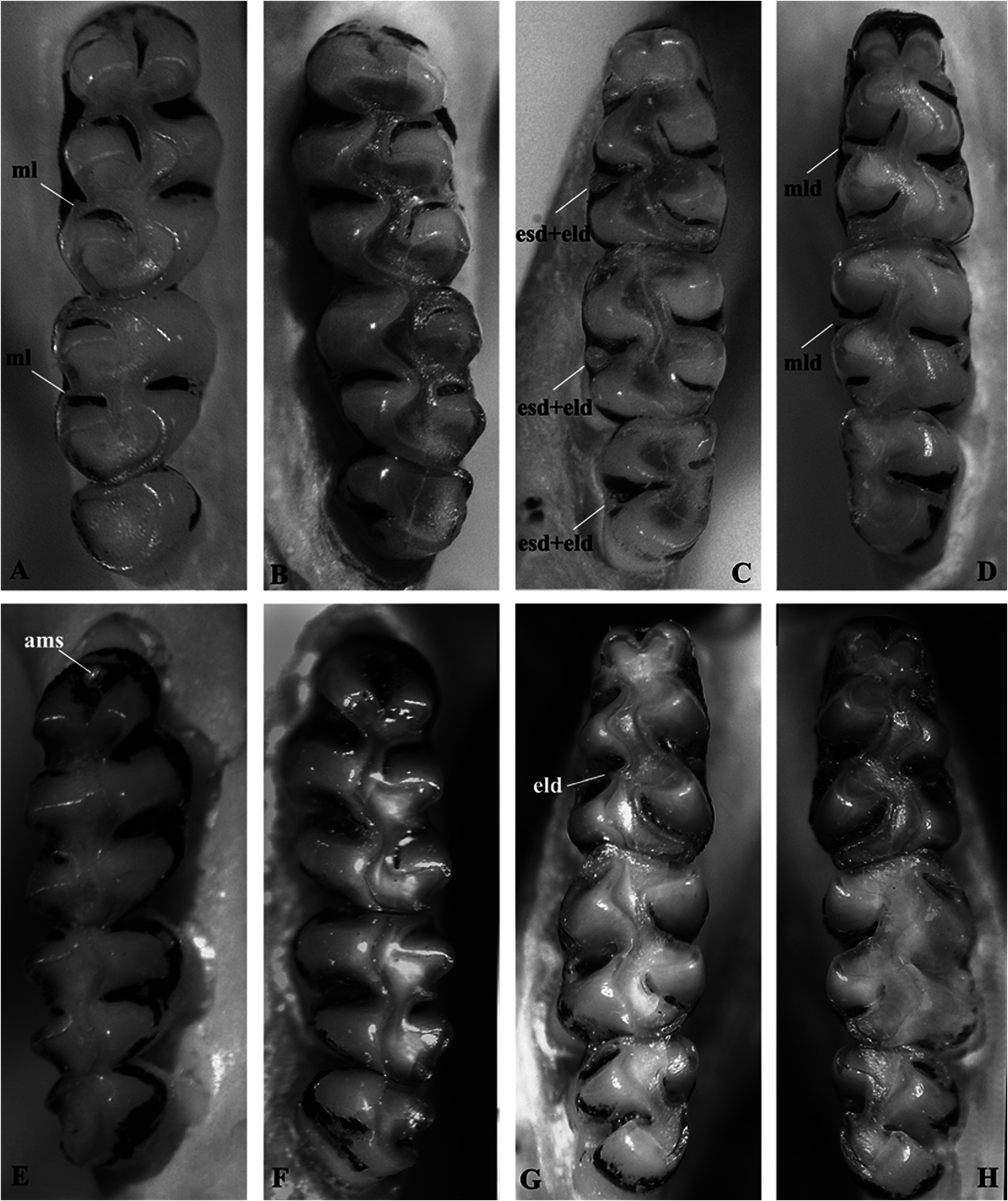
Upper and lower incisors slightly opisthodont, without grooves, and with enamel light orange. Upper molar rows parallel; main cusps arranged in slightly alternated pairs, with protocone and hypocone slightly anterior to opposite cusps (paracone and metacone, respectively); labial (paracone and metacone) cusps slightly larger than the lingual (protocone and hypocone) cusps; molars crested, narrow (M1 width mean: 1.2 mm) and elongated (molar toothrow length mean: 4.42 mm); upper and lower molars moderately hypsodont ( Fig. 8 View Fig ). M1 with conspicuous protoflexus, hypoflexus, paraflexus, and metaflexus, and inconspicuous anteroflexus and mesoflexus (obliterated with minimal wear); M1 separated by deep anteromedian flexus, resulting in an anterolabial conule subequal to or slightly larger than the anterolingual conule; anteroloph and parastyle present, but anteroloph coalesced to the anterolabial conule (thus, the anteroflexus is almost indistinguishable); protostyle absent; mesoloph reduced in length and generally coalesced with paracone due to wear ( Fig. 8A View Fig ); enteroloph and enterostyle absent; posteroloph poorly defined, probably fused to metacone due to wear (young specimens are unknown), with no evidence of posteroflexus. M2 similar to M1, rectangular in outline, with anteroloph moderately developed and fused to paracone at distal end, mesoloph very reduced and partially fused to paracone and metacone, flexus are less-developed, with no trace of a protoflexus. M3 half the size of M2, cylindrical in outline, with main cusps and flexus fused, hypoflexus inconspicuous; M3 generally distinctly worn ( Fig. 8B View Fig ).
Lower molars crested; labial cusps (protoconid and hypoconid) arranged in alternated pairs in relation to lingual (metaconid and entoconid) cusps, with metaconid and entoconid distinctly anterior to protoconid and hypoconid, respectively; m1 with anterolabial and anterolingual conulids of equal size, separated by a consistently present and deep anteromedian flexid; m1 with conspicuous protoflexid, hypoflexid, posteroflexid, and mesoflexid, and inconspicuous metaflexid and entoflexid; protolophid present, connected to anterior murid by enamel lophid and fused with protostylid; anterolophid and metastylid absent; mesolophid reduced (partially coalesced to entoconid due to wear) and fused to mesostylid ( Fig. 8D View Fig ); ectolophid present as narrow and slender structure or partially coalesced to hypoconid due to wear; ectostylid fused to ectolophid, but notably conspicuous ( Fig. 8C View Fig ); posterolophid conspicuous and separated from entoconid by well-defined posteroflexid; m2 similar to m1, with protolophid partially fused with protoconid due to wear; flexid less-developed, but distinct (except for a poorly developed protoflexid); m3 greater in size than M3, with the four main cusps visible, but opposite pairs fused due to wear; mesolophid and posterolophid might be present, but wear probably fused these structures with other cusps; flexids mostly fused, except for the hypoflexid; protoflexid and entoflexid inconspicuous (see specimen MZUSP 35766; Fig. 8C View Fig ).
Karyological data.— Chromosomal formula 2n = 40, FN = 40 ( Fig. 3 View Fig ).
Molecular divergence.— Cytochrome b haplotypes of A. kadiweu diverge, on average (p- distance), from those of A. philipmyersi , A. montensis , and A. toba by 7.6%, 8.0%, and 11.2%, respectively.
Ip, interparietal; la, lacrimal; lpp, lateral projection of the parietal; mas, mastoid; Max, maxillary; mf, mental foramen; mpp, median palatine process; ms, mesopterygoid fossa; Na, Nasal; nc, nasolacrimal capsule; Pa, parietal; Pal, palate; palc, posterior opening of the alisphenoid canal; pgf, postglenoid foramen; Pre, premaxillae; pt, parapterygoid fossa; sag, squamosal-alisphenoid groove; sn, sigmoid notch; spv, sphenopalatine vacuities; Sq, squamosal; ssf, subsquamosal fenestra; stf, stapedial foramen; szr, squamosal zygomatic root; zn, zygomatic notch; zp, zygomatic plate. White-dotted lines indicate relative position of some anatomical features.
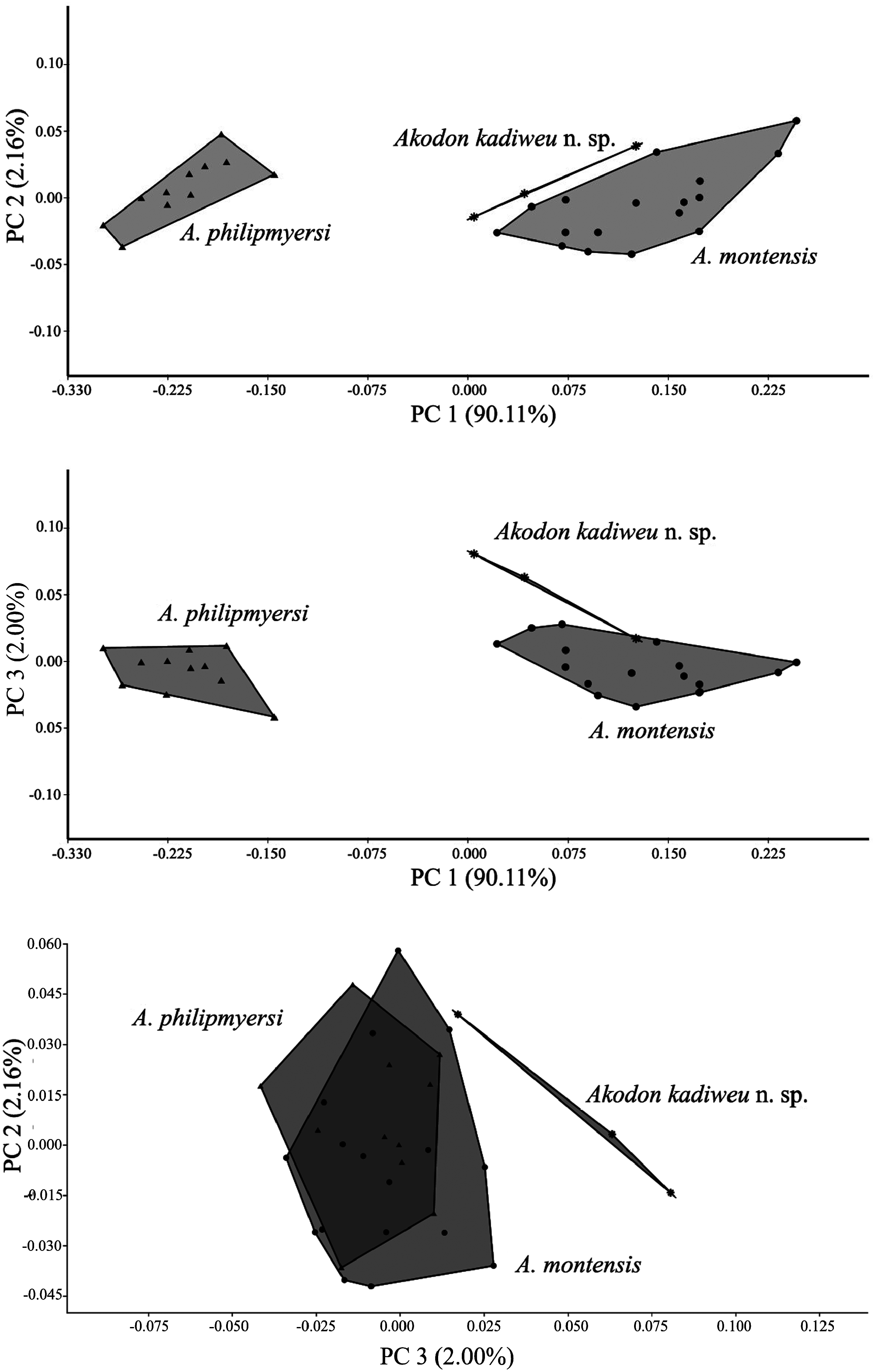
Comparisons.— Specimens of A. kadiweu exhibit larger values on all mean craniodental measurements analyzed than its sister taxon A. philipmyersi . In addition, these two species overlap only in four of the 15 skull measurements analyzed: OL, ZB, ZP, and BM1 (Table 2). Considering its greater overall size, A. kadiweu presents a relative narrower rostrum, as well as narrower zygomatic arches and zygomatic plates, compared to A. philipmyersi . The mean difference between values of zygomatic breadth (ZB) and braincase breadth (BB) is greater for A. philipmyersi (1.27 mm) than for A. kadiweu (0.94 mm), and can be clearly observed at side-by-side comparisons ( Fig. 6 View Fig ). Another distinctive feature is the longer tail of A. kadiweu (mean of 92% of HBL) compared to the distinctively shorter tail of A. philipmyersi (mean of 40% of HBL).
The pelage coloration also is very distinctive between these species, with A. philipmyersi exhibiting more defined limits between dorsal and ventral pelage coloration, while in A. kadiweu such transition is nearly undefined ( Fig. 5 View Fig ). Although A. kadiweu and A. philipmyersi present similar color at the base of dorsal fur (gray-based hairs in both species), the tips of setiform hairs are dark grayish-brown in the former and light brown (somewhat goldish-brown) in the latter. The ventral pelage of A. philipmyersi is gray-based with whitish/grayish-brown tips, while the venter of A. kadiweu is homogeneously dark gray, with only a brownish tip restricted to distal 1–2 mm of the hair (only visible under magnification). The overall body coloration of A. kadiweu is much darker than of A. philipmyersi , both in dorsal and ventral views ( Fig. 5 View Fig ). The mystacial vibrissae are denser and longer in A. kadiweu , reaching pinnae when laid back, in comparison to the short vibrissae of A. philipmyersi , which do not reach the pinnae.
The profiles of the skulls are notably different, as in A. philipmyersi it is distinctly domed and in A. kadiweu is flatter ( Fig. 6 View Fig ). Akodon kadiweu is also distinguishable from
a Data from Pardiñas et al. (2005).
A. philipmyersi by a long and slender rostrum, versus a short and blunt one, with premaxillae restricted to a narrow lateral strip in dorsal view ( Fig. 9D View Fig ); a narrower zygomatic plate, versus a wider and variable in shape, but in both species not as developed as in other congeners ( Fig. 9B View Fig ); poorly developed capsular projections of nasolacrimal foramina, versus well-developed projections ( Fig. 9D View Fig ); rostrum flanked by shallow zygomatic notches, versus wider and deeper notches ( Fig. 9D View Fig ); mesopterygoid fossa narrower than the parapterygoid fossae, versus fossae subequal in size ( Fig. 9E View Fig ); incisive foramina with posterior margins near the anterior protocone level of M1, versus posterior margins distinctly surpassing M1 protocone, near hypoflexus level ( Fig. 9A View Fig ); palate short, with anterior margin of mesopterygoid fossa aligned with posterior margin of M3, versus a long palate, with anterior margin well posterior to the molars ( Fig. 9E View Fig ); frontosquamosal suture notably anterior to frontoparietal suture, which allows contact between the frontal and the dorsal surface of squamosal, versus frontosquamosal suture only slightly anterior to the frontoparietal suture, with frontal and dorsal surface of squamosal not or only slightly in contact ( Fig. 9C View Fig ). The mandibles of both species are very similar, but more delicate in A. philipmyersi ( Fig. 6 View Fig ).
General molar morphology is similar between A. kadiweu and A. philipmyersi , except that all cusps are somewhat more compressed in A. philipmyersi , particularly in upper molars ( Fig. 8 View Fig ). Both species present anterolophs, mesoloph/ids, ectolophids, and ectostylids. Nonetheless, such structures are more developed in A. kadiweu , which presents more developed anterolophs and mesolophs (but both fused to adjacent cones due to wear); mesolophid of m1 and m2 small, but distinct (fused to mesostylid); ectolophid fused to ectostylid in m1, m2, and m3, forming a conspicuous structure in m1 and m2. In A. philipmyersi , comparatively, the anteroloph and mesoloph are less-developed; the mesolophid of m1 is generally smaller and in m2 is vestigial (or absent), both very reduced and frequently not fused to mesostylid (when present); ectolophid very reduced in m1 and m2 (absent in m3) and generally not fused to ectostylid (when present not conspicuous). Moreover, few specimens of A. philipmyersi present an anteromedian style ( Fig. 8E View Fig ; also see Barbière et al. 2019: figure 5), a structure not seen in A. kadiweu (or any other specimen of the genus Akodon here analyzed).
We also provide further comparisons of A. kadiweu to congeners recorded in the vicinities of Serra da Bodoquena ( Fig. 10 View Fig ), namely A. montensis and A. toba (see Pardiñas et al. 2015). Both A. kadiweu and A. montensis present brown dorsal pelage, but the former is much darker; the ventral pelage of A. montensis is gray-based with yellowish tips, while the venter of A. kadiweu is homogeneously dark gray, with only a brownish tip restricted to distal 1–2 mm of the hair (only visible under magnification) ( Fig. 5 View Fig ). Both species are morphometrically similar, greatly overlapping in all body and cranial dimensions analyzed herein, but A. montensis is larger in most measurements means, except for IOC, BB, MTRL, and OCW (Table 2). Both species can be distinguished by form, not only size (see the results of PCA above; Fig. 4 View Fig ). There also are some cranial characters that distinguish A. kadiweu and A. montensis . Both species present hourglass-shaped interorbital region, but the interorbital constriction is more pronounced in A. montensis than in A. kadiweu ( Fig. 6 View Fig ), in which the lateral margins of the supraorbital region converge to the interorbital constriction in a nearly straight line (most similar to A. philipmyersi ; see also Gonçalves et al. 2007:6); the incisive foramina are longer in A. montensis , nearly reaching the posterior end of the protocone of M1, while shorter in A. kadiweu , only reaching the anterior margin of protocone of M1; the zygomatic plate is distinctly more developed in A. montensis ( Fig. 6 View Fig ). Only A. kadiweu present a prominent median palatine process in the anterior border of the mesopterygoid fossa while in A. montensis there is no trace of a such structure, exhibiting an “inverted U-shaped” mesopterygoid fossa; A. kadiweu exhibits a projection of parietal onto the squamosal ( Fig. 7F View Fig ), and A. montensis a straight lateral border of parietal ( Fig. 6 View Fig ). Dental characters in lower molars allow distinction between both species, as the ectolophid fused to ectostylid form a conspicuous structure in A. kadiweu ( Fig. 8C View Fig ), but in A. montensis both structures are commonly absent or, when rarely present, never as developed as in A. kadiweu .
Besides the conspicuous larger size (CIL mean = 27.30 mm, MTRL mean = 4.70 mm —Myers 1990), A. toba also is qualitatively very distinctive to A. kadiweu . Dorsal pelage of A. toba is strongly olivaceous, with an eye-ring consistently present, and the ventral pelage is paler, grayish frosted or buffy (strongly contrasting with dorsum), very distinct to the overall much darker pelage of A. kadiweu , which has no trace of eye-ring, except for MZUSP 35994 (Supplementary Data SD3), which is not as conspicuous as in A. toba . The skull and mandible of A. toba are more heavily built, with proportionally shorter rostrum (RL/CIL 37% mean), broad and deep zygomatic notches, zygomatic arches widely flared, zygomatic plates notably broad, interorbital region with squared or beaded edges, supraorbital and temporal ridges well-developed, narrower mesopterygoid and parapterygoid fossae, both with straight borders, well-developed mastoid ridges, and parietal notably expanded onto lateral surface of braincase (posteriorly to the end of the squamosal zygomatic root; see Weskler 2006: figure 15B).
All species that occur in Mato Grosso do Sul ( A. toba and A. montensis ), as well as its sister taxon A. philipmyersi , exhibit proportionally shorter tails (TL/HBL ratio averages 42%, 81%, 40%, respectively—Myers 1990; Pardiñas et al. 2005, 2015) compared to A. kadiweu . The average of 92% of TL/HBL ratio is the most distinctive external morphological feature of A. kadiweu and no other species of Akodon exhibits this tail/ body length proportion (see Pardiñas et al. 2015). Allied to the 2n = 40, FN = 40 karyotype and the dark overall body pelage coloration (grayish-brown), the traits described above mostly distinguish this new species of Akodon from its congeners.
Natural history.— Specimens of A. kadiweu were collected in a fragment of deciduous dry forest. Specimens were captured during the end of wet season, in April, three with Sherman (7.5 × 8.5 × 23 cm) and one with Young (19.5 × 20 × 32 cm) live traps, in ground level. Traps were baited with a mixture of peanut butter, sardines in oil, and corn meal, fixed on a slice of manioc. Individuals showed no trace of reproductive activity.
Other small mammals that were collected in the same deciduous dry forest fragment (by the senior author, during the same field trip in April 2002) were: Marmosa rapposa (sensu Voss et al. 2020), Monodelphis domestica , Hylaeamys megacephalus , Oecomys franciscorum , Oligoryzomys chacoensis , Ol. nigripes , and Thrichomys fosteri , revealing a biogeographical affinity with small mammal assemblages from the Chaco ( Carmignotto et al. 2012).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.