
Aglaophenopsis cornuta (Verrill, 1879)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3737.5.1 |
|
publication LSID |
lsid:zoobank.org:pub:B5FE322D-4D0A-45E6-84BF-F00FA6308DE1 |
|
DOI |
https://doi.org/10.5281/zenodo.6149342 |
|
persistent identifier |
https://treatment.plazi.org/id/03AEB111-0A4E-BD20-99AC-FA07FBF37911 |
|
treatment provided by |
Plazi |
|
scientific name |
Aglaophenopsis cornuta (Verrill, 1879) |
| status |
|
Aglaophenopsis cornuta (Verrill, 1879) View in CoL
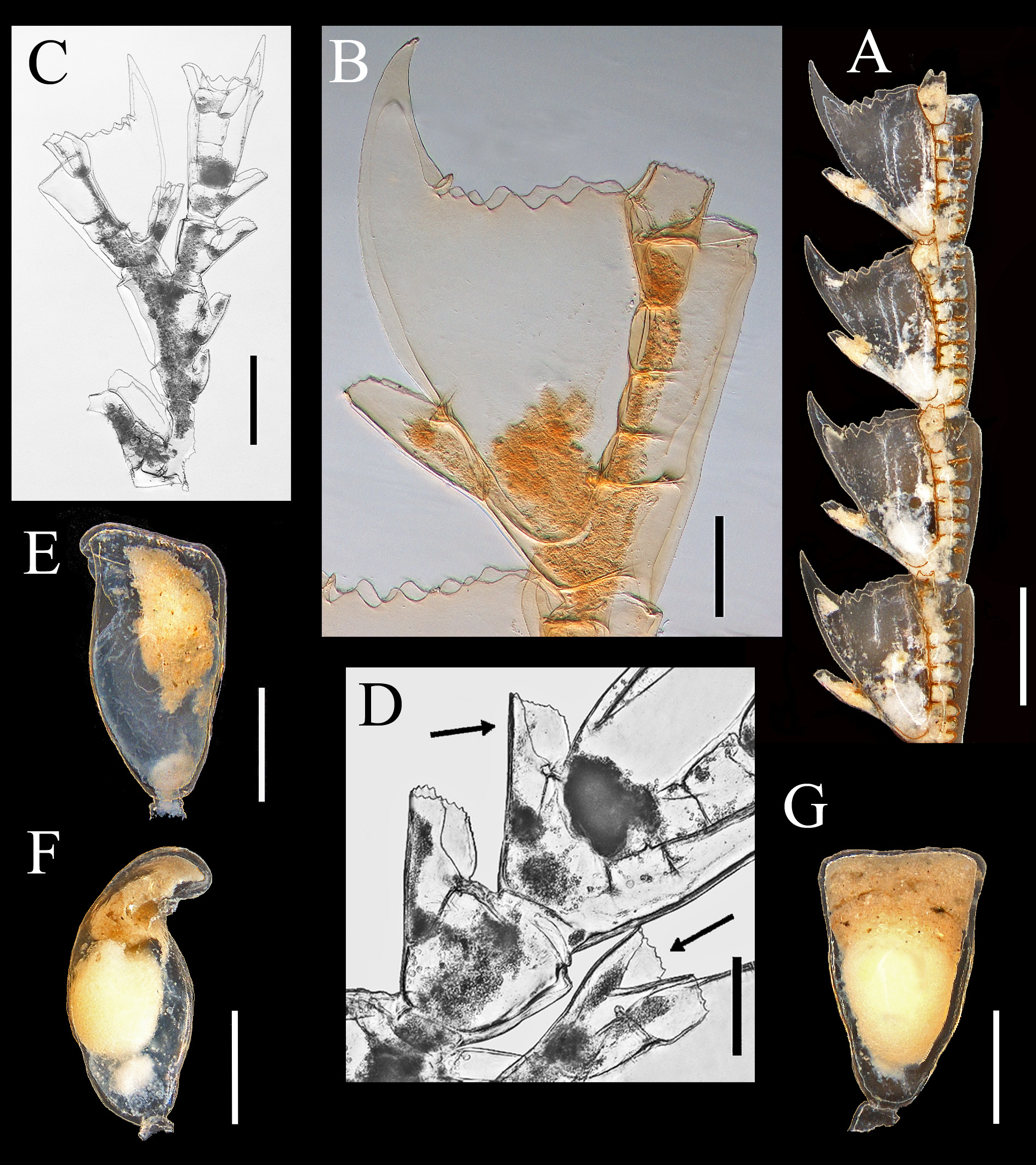
( Figs. 1 View FIGURE 1 A, 2A–G, tables 1, 3, 9–12)
Cladocarpus cornutus Verrill, 1879: 310 .—Levinsen 1893: 208, pl. 8, fig. 19–22.—Vanhöffen 1897: 246.—Jäderholm 1909: 110.—Kramp 1913: 28.—Kramp 1914: 1059.—Fraser 1946: 417.—Calder 1970: 1540, pl. 8, fig. 7.—Peña Cantero & García Carrascosa 1999: 214.—Schuchert 2001: 137, fig. 117A–E.—Bouillon et al. 2006: 283.
Aglaophenia cornutus: Bonnevie 1899: 94 .
Aglaophenopsis cornuta: Nutting 1900: 120 , pl. 30, fig. 6–9.—Whiteaves 1901: 28.—Broch 1910: 234.—Kindle & Whittaker 1917: 232.—Broch 1918: 77, fig. 39a–b, fig. 40a–d, fig. 41.—Jäderholm 1919: 11.—Fraser 1921: 177, fig. 102.—Kramp 1932a: 56.—Kramp 1932b: 20.—Kramp 1943: 44.—Fraser 1944: 395, pl. 85, fig. 382.—Vervoort 1972: 204.—Calder 1997b: 88.—Henry 2001: 163.
Material examined. FN3L06 L51, 2 fertile colonies, largest one 9.5 cm high; FN3L06 L85, one fertile colony 7.0 cm high; FN3L06 L88, one fertile colony 10.5 cm high; FN3L06 L92, one fertile colony 15.0 cm high; PLA07 L61, one sterile colony 8.4 cm high; PLA07 L99, one sterile colony 10.0 cm high; PLA07 L107, one fertile colony 6 cm high; FC07 L74, one sterile colony 7.2 cm high; FC07 L133, one sterile colony 4.3 cm high; FC07 L153, one sterile colony 13.0 cm high; FC07 L171, one sterile fragment 1.6 cm high; FC07 L177, one sterile colony 5.5 cm high; FC07 L178, one fertile fragment 6.2 cm high; FN3L08 L88, 3 fertile colonies, largest one 21.0 cm high; FN3L08 L89, 3 fertile colonies, largest one 15.0 cm high; FN3L09 L80, one fertile colony 10.0 cm high; FN3L09 L89, one fertile colony 16.0 cm high; FN3L10 L35, one fertile colony 14.0 cm high; FN3L10 L57, one fertile colony 9.0 cm high; FN3L10 L87, one fertile colony 15.5 cm high; NEREIDA0509 RD4, one fertile colony 12.2 cm high; NEREIDA0509 RD6, 5 colonies, largest one 15.3 cm high, 2 of them fertile; NEREIDA0509 RD7, 6 colonies, up to 15.6 cm high, some fertile; NEREIDA0609 RD26, 2 sterile colonies, largest one 4.2 cm high; NEREIDA0609 RD27, 3 colonies, largest one 5.5 cm high, one fertile; NEREIDA0609 RD28, 2 fertile colonies, both 6.0 cm high; NEREIDA0609 RD41, 34 colonies, up to 11.3 cm high, some fertile; NEREIDA0709 RD49, 5 colonies, largest one 6 cm high, 2 fertile; NEREIDA0709 RD59, 2 fertile colonies, largest one 9.5 cm high; NEREIDA0709 RD60, one sterile colony 5.2 cm high; NEREIDA0610 RD62, one sterile colony 2.0 cm high; NEREIDA0610 RD67, one fertile colony 3.0 cm high; NEREIDA0610 RD71, one sterile fragment 2.2 cm high; NEREIDA0610 RD74, 23 colonies, up to 10.3 cm high, some fertile; NEREIDA0710 RD75, one sterile colony 6.0 cm high; NEREIDA0710 RD76, one fertile colony 8.7 cm high; NEREIDA0710 RD77, 2 sterile colonies, largest one 5.7 cm high; NEREIDA0710 RD78, one sterile colony 3.0 cm high; NEREIDA0710 RD79, one fertile colony 10.2 cm high; NEREIDA0710 RD80, 8 colonies, up to 8.4 cm high, some fertile; NEREIDA0710 RD87, one fertile colony 12.0 cm high; NEREIDA0710 RD88, 3 colonies, largest one 6.6 cm high, 2 fertile; NEREIDA0710 RD92, 2 sterile colonies, largest one 8.4 cm high; NEREIDA0810 RD97, 5 colonies, largest one 8.0 cm high, 3 fertile; NEREIDA0810 RD98, 4 colonies, largest one 8.4 cm high, one fertile; NEREIDA0810 RD99, 13 colonies, up to 8.5 cm high, some fertile; NEREIDA0810 RD100, 3 sterile colonies, largest one 9.0 cm high; NEREIDA0810 RD101, one sterile colony 9.1 cm high; NEREIDA0810 RD102, 2 colonies, largest one 7.8 cm high, one fertile; NEREIDA0810 RD103, 9 colonies, up to 18 cm high, some fertile; NEREIDA0810 RD104, 15 colonies, up to 28.7 cm high, some fertile.
Description. Colonies up to 28.7 cm high, formed by a broad hydrorhizal mass supporting a polysiphonic stem, up to 3.0 mm wide proximally, thinning out to monosiphonic distally. Stem rigid, thick, erect. Ramification in one plane, profuse, up to the fourth order. Branches almost opposite, arising at slightly acute angles, long, polysiphonic for most of their length, slightly tapering distally. Stem deep brown basally and light brown distally; branches also light brown.
Main axial tube on front of stem and branches, divided into short internodes, 550–600 µm long. Successive internodes each with one apophysis alternately right and left, giving rise to branches and hydrocladia. Three nematothecae surrounding the apophyses: one basal and two distal. Accessory tubes numerous, with neither hydrothecae nor nematothecae.
Hydrocladia alternate, up to 1.2 cm long, divided into up to 15 thecate internodes by more or less well-marked transverse nodes. Internodes with 2–9 internal septa, one hydrotheca, and three one-chambered nematothecae. Hydrotheca as long as the internode, large, conical in lateral view and somewhat oval to round in cross-section, with depth/width at rim= 1.08–1.48 (n= 25). Intrathecal septum inconspicuous, very small, arising from the adaxial wall of hydrotheca and projecting forwards in its lumen. A characteristic abcauline median keel with a sharp, hollow tip, arising above mesial nematotheca; strong, large, laterally compressed, slightly curved, extending 332– 528 µm beyond level of hydrothecal margin, reaching end of mesial nematotheca of the following distal internode, or nearly so. Hydrothecal rim with 9–11 low cusps, including a small mesial tooth aligned with the keel.
Mesial nematothecae extending for 1/3–1/2 the length of abaxial hydrothecal wall, slightly tapering distally, with wide aperture and crenulated rim, as well as a thickening of perisarc at its basis in the axil between the wall of the hydrotheca and the nematotheca; on first internode it is slightly displaced to one side. Lateral nematothecae arising from hydrocladium, partially attached to distal hydrothecal wall, with end slightly surpassing hydrothecal rim and crenulated margin.
Phylactocarps forked ( Y –shaped), segmented, up to 2.3 mm long, arising laterally from hydrothecal bases of proximal thecate internodes beside mesial nematotheca; not replacing nematotheca. Internal septa numerous. Segments short, carrying unpaired nematothecae with margin of aperture finely crenulated. Both branches with one terminal hydrotheca. Gonothecae borne on short pedicels, obovate, curved, arising singly from apophyses of stem and branches; aperture broadly oval to almost circular, latero-distal. Sex could not be ascertained.
Remarks. This species is readily distinguished from others in the study area by the characteristic shape of its hydrothecae ( Figure 2 View FIGURE 2 A–C). Especially noteworthy is the presence of a pronounced median hydrothecal keel with thickened perisarc. Its tip normally reaches the base of the mesial nematotheca of the following internode, and sometimes also approaches its end. The variability of this keel was discussed by Broch (1918). Aglaophenopsis bonnevieae (Jäderholm, 1909) , sometimes sympatric with A. cornuta in Greenland and subarctic waters of the northwestern Atlantic (see Kramp 1932a; Schuchert 2001), likewise has a prominent, although somewhat less developed, hydrothecal keel. Moreover, its hydrotheca differs from that of A. cornuta in having a conspicuous intrathecal septum, and the hydrothecal rim is slightly crenulated rather than cusped. Paired nematothecae on the internodes of the main axial tube also occur in the upper axil of the apophyses rather than at its sides. Phylactocarps of A. bonnevieae also differ in being much more ramified and in having more hydrothecae.
Hydrothecae of A. cornuta somewhat resemble certain species of Gymnangium Hincks, 1874 , including the European G. montagui (Billard, 1912) and others from South Africa (see Millard 1975). However, the ‘horn’ in such species of Gymnangium is a hypertrophied mesial nematotheca, while that of A. cornuta is a large median hydrothecal keel arising distal to the mesial nematotheca. The keel does not represent a highly modified median abcaulinar cusp of the hydrothecal margin because that cusp is both present and small.
The margin of the median inferior nematotheca of the thecate internodes (and also of the terminal hydrothecae of the phylactocarp), is at times deeply indented in the middle, giving the impression of a twin theca (NEREIDA0710 RD79, Figure 2 View FIGURE 2 D).
Colonies with gonothecae on stem apophyses, unprotected by any phylactocarps, were found in samples from some stations (NEREIDA0709 RD49).
Although originally assigned to Cladocarpus , this species has also been referred to Aglaophenopsis by several authors. According to Fraser (1944), phylactocarps of this species represent a modified mesial nematophore. In our material, however, the phylactocarp arises laterally from the base of a hydrotheca borne on a proximal thecate internode. It is situated beside a clearly distinct mesial nematotheca and does not replace it. Moreover, contrary to Fraser, both branches of the phylactocarp bear a terminal hydrotheca. The species was assigned to Cladocarpus (as C. cornutus ) by both Bouillon et al. (2006) and Schuchert (2012a).
Observed depth range: 240–1885 m. The known maximum depth for the species is extended from 1200 m (Kramp 1932a) to 1885 m. A neritic-bathyal species (Calder 1997b).
Fertile material. Collected in August 2006 (240–1233 m depth), June and July 2007 (460–878 m depth), August 2008 (392–1198 m depth), from May to August 2009 (425–1885 m depth), and from June to August 2010 (345–1353 m depth).
Distribution. A North Atlantic species known from Denmark Strait (Broch 1918, ca. 253–603 m; Schuchert 2001, 200– 700 m), eastern Greenland (Broch 1918, ca. 260–621), southwest Greenland at 63º36’N – 55º15’W (1200 m) and 60º16’N – 47º48’W (135 m) (Kramp 1932a), two stations in Bredefjord, south Greenland (Kramp 1932b, 310–330 m and 700 m), Davis Strait (Levinsen 1893, ca. 188–544 m; Kramp 1913, 700– 1055 m; Broch 1918, ca. 544 m), western Greenland (Broch 1918, ca. 1095 m), Baffin Bay (Jäderholm 1909, ca. 731 m), 72°32’N – 58°05’W (ca. 212 m), 60º27’N – 48º31’W (Vervoort 1972, 326– 366 m), 53°34’N – 52°01’W (ca. 1792 m) and 52°25’N – 52°12’W (ca. 296 m) (Jäderholm 1919), eastern Canada (Kindle & Whittaker 1917, ca. 365 m, based on literature records), off northeast Newfoundland, 51°22’30”N – 50°31’30”W (Calder 1970, 457 m, as Cladocarpus cornutus ), Flemish Cap, Flemish Pass, and Grand Banks (present study), Northeast Channel, south of Nova Scotia, at 42°02’N – 65°36’W (Henry 2001; personal communication, 24.11.2012), off Sable Island on Banquereau (Verrill 1879, ca. 365 m, first description; Nutting 1900; Whiteaves 1901; Fraser 1918, 1944), east of Gloucester (Massachusetts) (Fraser 1944, ca. 310 m). Not recorded further south by Henry et al. (2008), but there is a recent southern record from 53º56’S – 63º51’W (250 m) by El Beshbeeshy (2011).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |