
Acestrorhynchus undetermined
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2007.00355.x |
|
persistent identifier |
https://treatment.plazi.org/id/5C71FF4E-3275-3D3C-4480-FF69E62CFF34 |
|
treatment provided by |
Felipe |
|
scientific name |
Acestrorhynchus undetermined |
| status |
|
PHYLOGENY OF ACESTRORHYNCHUS SPECIES 731
posterior portion of the ischiatic process is herein proposed as an autapomorphy for A. minimus .
85. Bony lamella on lateral side of pelvic bone (0, extending along up to half of lateral margin; 1, reaching to anterior tip of pelvic bone) (CI = 100; RI = 100).
The shape of the pelvic bone of the species of Acestrorhynchus is similar to that described by Weitzman for B. meeki (1962: 42, fig. 21), and consists of a rod-shaped element with a flat bony lamella along its medial margin that gradually tapers anteriorly. Lateral to the rod-shaped element there is a similar, but less-developed, bony lamella that extends anteriorly up to approximately half of the length of the pelvic bone.
In Cynopotamus , Charax , Roeboides , and Acanthocharax the bony lamella situated lateral to the rodshaped element is more developed than in the condition described for Acestrorhynchus , and extends to the anterior tip of the pelvic bone and is similar in shape to the lamella of the medial side of the pelvic bone. As a consequence, the pelvic bone in the former three genera is more symmetrically shaped, with the rod-shaped element situated in the centre of the ossification instead of having a more lateral position. Although the degree of development of the bony lamella lateral to the rod-shaped element may vary among other characiform outgroups, it is never as developed as the medial bony lamella. Furthermore, it usually does not extend beyond half of the extension of the rod-shaped element.
86. Bony lamella on medial side of pelvic bone (0, extending up to anterior tip of pelvic bone; 1, reduced or extending up to approximately half the length of pelvic bone) (CI = 100; RI = 100).
The flat bony lamella medial to the rod-shaped element of the pelvic bone is usually present, although variously developed among characiforms, and extends to, or almost to, the anterior limit of the rod-shaped element (state 0). The lamella is widest posteriorly and tapers anteriorly. In Cynodon and Rhaphiodon this lamella is reduced overall and is evident only at the posterior portion of the pelvic bone, and does not extend beyond the middle of the length of that ossification (state 1). All cynodontines possess relatively elongate pelvic bones, but in the species of Hydrolycus a narrow bony plate is still present along the medial margin of the rod-shaped element. Although the reduction of the medial bony lamella is more pronounced in Rhaphiodon than in Cynodon , in both genera most of the pelvic bone is represented exclusively by the long rod-shaped element. The reduced bony lamella on the medial side of the pelvic bone is proposed as an additional synapomorphy of Cynodon and Rhaphiodon ( Toledo-Piza, 2000) .
87. Presence, absence, and degree of development of anteroventral projection on ischiatic process (0, absent or reduced; 1, well developed) (CI = 50; RI = 66).
The presence of a long anteroventral projection on the ischiatic process has been proposed as a synapomorphy for a clade comprising Phenacogaster , the Cynopotaminae (sensu Menezes, 1976), Acanthocharax , Charax , and Roeboides (Lucena, 1998) . In the present study a similar process was observed in Astyanax , Moenkhausia , and Tetragonopterus . In some species of Acestrorhynchus (e.g. A. altus , A. britskii , A. falcatus , and A. pantaneiro ) there is a very short projection ventrally on the ischiatic process; however, the projection is never as developed as the process present in the aforementioned taxa, and in addition it is somewhat variable inter- and intraespecifically. Therefore, the condition in Acestrorhynchus was coded as state 0.
ANAL FIN
88. Process on proximal radial of seventh anal-fin pterygiophore (0, absent; 1, present) (CI = 50; RI = 83).
All species of Acestrorhynchus except A. abbreviatus , A. altus , A. falcatus , A. heterolepis , A. lacustris , and A. pantaneiro , possess a posteriorly directed process on each side of the distal portion of the proximal radial of the seventh anal-fin pterygiophore (state 1; Fig. 22A View Figure 22 ). A weak band of ligamentous tissue is attached to the tip of the process of the seventh anal-fin pterygiophore, and it extends to the anal-fin stay where it also attaches. This ligamentous band also attaches to the seventh anal-fin pterygiophore in A. abbreviatus , A. altus , A. heterolepis , A. falcatus , A. lacustris , and A. pantaneiro : species in which the process is lacking (state 0; Fig. 22B View Figure 22 ). In these last six species there is a slight sinuosity in the region occupied by the process in the remaining species of Acestrorhynchus . It was not possible to clearly determine the presence/absence of this ligamentous stripe and its site of attachment in outgroup taxa; however, an ossified process on the seventh pterygiophore is absent in all examined outgroups. Under hypothesis 1 the interpretation of the presence of this process is, however, ambiguous, and using the ACCTRAN optimization procedure it is most parsimoniously interpreted as a synapomorphy of Acestrorhynchus , with a loss of this process in the clade formed by A. abbreviatus , A. altus , A. heterolepis , A. falcatus , A. lacustris , and A. pantaneiro . Under hypothesis 2 the presence of an ossified process on the seventh anal-fin pterygiophore is interpreted as independently acquired in the clade formed by A. britskii , A. grandoculis , A. microlepis , and A. minimus , and the clade formed by A. falcirostris , A. isalineae , and A. nasutus .
89. Medial bony lamella on proximal radial of first anal-fin pterygiophore (0, present; 1, absent) (CI = 25; RI = 50).
The proximal radial of the first anal-fin pterygiophore in all species of Acestrorhynchus except A. grandoculis , A. microlepis , and A. minimus has a median bony lamella located distally along the anterior margin of the ossification ( Fig. 22B View Figure 22 ): a condition similar to that of B. meeki ( Weitzman, 1962: fig. 17a). In the latter three species of Acestrorhynchus such a median lamella is absent ( Fig. 22A View Figure 22 ), and in anterior view the distal anterior portion of the proximal radial of the first anal-fin pterygiophore has two short ridges that are continuous with the anterior lateral and medial margins of the remaining portion of the first pterygiophore. Under this condition, only the distalmost portion of the first pterygiophore is slightly expanded relative to the rest of the ossification, with the anteriormost unbranched anal-fin rays attaching to this expanded portion of the bone.
A median bony lamella is present in all examined characiforms except Boulengerella , Hepsetus , Hoplias , Hoplerythrinus , and Hydrolycus . In all these taxa, however, the distal portion of the first anal-fin pterygiophore is not expanded, thereby differing in shape from the distal portion in the Acestrorhynchus species that lack a median bony lamella. Despite this difference in these bones, all these taxa were coded as lacking a medial bony lamella on the proximal radial of the first anal-fin pterygiophore. The lack of this lamella is proposed as a synapomorphy for the clade formed by A. grandoculis , A. microlepis , and A. minimus under both of the hypotheses derived from the present analysis.
CAUDAL FIN
90. Number of epurals (0, three; 1, two; 2, one) (ordered) (CI = 66; RI = 0).
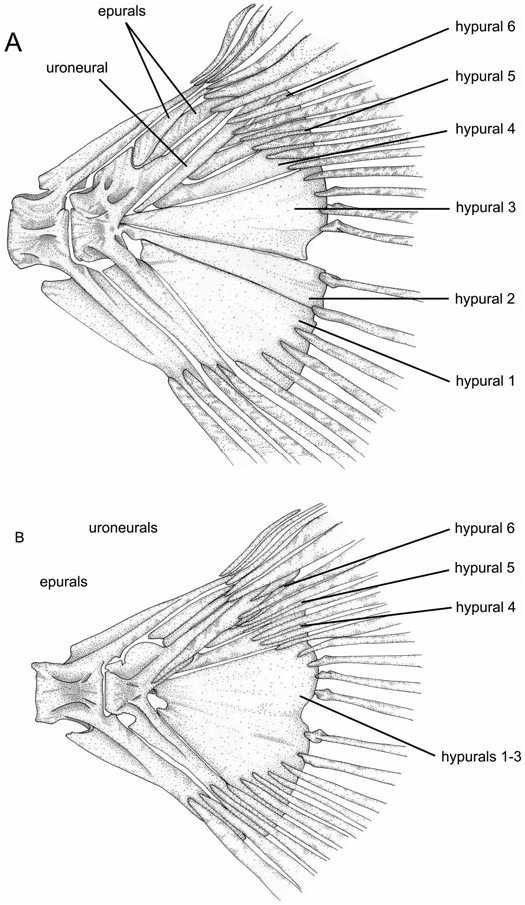
Epurals are laterally compressed bony elements situated on the dorsal portion of the dorsal-fin skeleton between the urostyle and last neural spine ( Weitzman, 1962: fig. 15). In characiforms the number of epurals varies from one to three ( Roberts, 1969: 428).
All species of Acestrorhynchus possess two epurals ( Fig. 23 View Figure 23 ). One specimen of A. nasutus (MZUSP 47698) has one epural, but its distal tip is bifurcated, and all other examined specimens of this species have two epurals, so therefore the condition for this species was coded as 1. All examined outgroups also possess two epurals except for Hepsetus and Boulengerella , with three, and Hoplias , with one. Although Hydrolycus tatauaia has three epurals, all other Hydrolycus species , Cynodon , and Rhaphiodon have two epurals. H. tatauaia was hypothesized to be more derived relative to the remaining species of Hydrolycus (Toledo- Piza, 2000), and the condition in Hydrolycus was therefore coded as 1.
91. Hypurals 2 and 3 (0, not in contact along their margins; 1, their margins contacting anteriorly, but separated posteriorly; 2, in contact along their entire margins; 3, fused). (ordered) (CI = 33; RI = 75).
Four different types of contact between hypurals 2 and 3 were observed among the examined taxa. In Charax , Boulengerella , Hoplias , and Moenkhausia these two elements are separated by a gap along most of their adjacent margins, in a manner similar to that illustrated by Roberts for Hepsetus , Ctenolucius , Hoplias , Salminus , and Hydrocynus ( Roberts, 1969: figs 56–60) (state 0). In Cynopotamus , Roeboides , Oligosarcus , and Xenocharax hypurals 2 and 3 are in contact anteriorly, but posteriorly their margins gradually diverge, leaving a space between these two ossifications (state 1). In Acestrorhynchus and the Cynodontinae there is no gap between the adjacent margins of hypurals 2 and 3, and these elements are either in close contact along their entire length (state 2) or are fused (state 3). A small notch is usually present at the posteroventral margin of hypural 3 (when the margins of hypurals 2 and 3 are not fused) or in the posterior margin formed by the fused hypurals 2 and 3, but this notch does not extend anteriorly to form a gap between the two elements, as in state 1. The fusion of the adjacent margins of hypurals 2 and 3 along most of their lengths was proposed as a synapomorphy for the Cynodontidae plus Acestrorhynchidae ( Lucena & Menezes, 1998) . These two elements are fused in all cynodontines with at least some degree of fusion already present in relatively small specimens ( Toledo-Piza, 2000: 50). In Acestrorhynchus there is both ontogenetic and interespecific variation in the degree of fusion between hypurals 2 and 3, and other features of the caudal fin in the genus were described by Miquelarena (1982: 288). In small specimens of all species, these two elements are autogenous (i.e. it is still possible to observe the limit between the two hypurals). In larger specimens of A. falcatus and A. falcirostris the suture between the hypurals is still present. These sutures are also evident in species that achieve only relatively small body sizes as adults, such as A. minimus and A. nasutus . In A. grandoculis there is some degree of fusion, with at least part of the adjoining edges between these bones not evident; the same condition being present in the remaining Acestrorhynchus species. Roestes and Gilbertolus have autogenous hypurals 2 and 3, and a small notch at the posteroventral portion of the latter element. The condition in these taxa is, however, more similar to that of Acestrorhynchus than to species coded as state 1. Therefore these species were also coded as state 2, in agreement with Lucena & Menezes (1998: 264). The condition of hypurals 2 and 3 in contact along their entire margins, but not fused to each other, is herein interpreted as synapomorphic for the clade formed by A. falcirostris , A. isalineae , and A. nasutus , with independent acquisitions in A. falcatus and A. minimus .
LATEROSENSORY CANAL SYSTEM IN

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |