
Aaptos kanuux, Lehnert & Hocevar & Stone, 2008
|
publication ID |
https://doi.org/ 10.11646/zootaxa.1939.1.8 |
|
persistent identifier |
https://treatment.plazi.org/id/3F2E1742-E056-FFE3-97E5-59AD2DF0A9B5 |
|
treatment provided by |
Felipe |
|
scientific name |
Aaptos kanuux |
| status |
sp. nov. |
Aaptos kanuux sp. n.
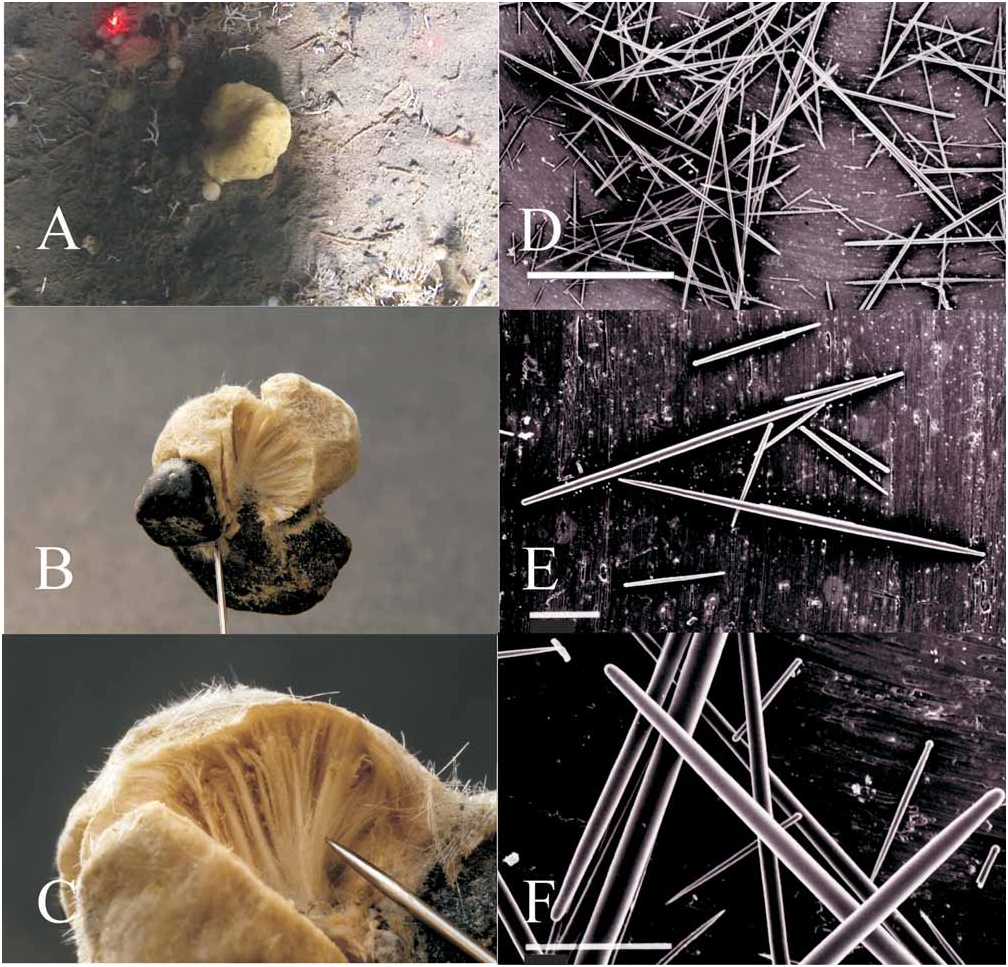
Holotype: USNM 1117764 About USNM ; Figs. 1A–C View FIGURE 1 , stored in ethanol after collection. Collected by Kenneth Lowyck in Pribilof Canyon at 219 m depth, 1 August 2007, 55°59.431N, 170°01.378W. GoogleMaps
Description. The sponge is irregularly globular, somewhat wider than high, diameters range from 9 to 19 mm ( Fig. 1B View FIGURE 1 ). At the base the sponge is firmly attached to two small black pebbles ( Fig. 1B View FIGURE 1 ). The surface is almost smooth, microscopically slightly uneven and in places microhispid, due to protruding spicules. Live color is mustard yellow ( Fig. 1A View FIGURE 1 ), in ethanol the sponge is greyish beige. In life, the black pebbles were buried in the silt with only the sponge protruding ( Fig. 1A View FIGURE 1 ). No oscules are visible, neither on the collected specimen, nor on videos. The consistency is firm, only slightly elastic, and almost incompressible. The radial arrangement of the spicules in the interior of the sponge is evident with the unaided eye on sections perpendicular to the surface ( Fig. 1C View FIGURE 1 ).
Spicules. Spicules are strictly arranged radially in all parts of the sponge ( Fig. 1C View FIGURE 1 ); spicule density is very high throughout the sponge and thus the consistency is hard. There is no cortex. Single spicules are grouped in polyspicular tracts which are slender at start (as paucispicular tracts, 30µm in diameter) containing only few spicules per cross section. These polyspicular tracts widen towards the surface and reach diameters of 420µm with approximately 20 spicules per cross section. Small tylostyles form a dense palisade at the surface. Spicules at the ends of the polyspicular tracts can protrude through the palisade in some places and cause the microhispid surface there. Three types of spicules are proper to the sponge ( Figs. 1 View FIGURE 1 D-F) and represent the typical set of spicules of the genus: ectosomal tylostyles ( Figs. 1 View FIGURE 1 E-F) measure 104 – 215 x 4-8µm; choanosomal spicules are strongyloxeas ( Fig. 1D View FIGURE 1 ), 1795 – 2132 x 15-22 µm and, fusiform subtylostyles ( Fig. 1E View FIGURE 1 ), occasionally strongyloxeas ( Fig. 1F View FIGURE 1 ), when the tyle is missing, 485 – 770 x 8-10µm.
Distribution: It is known only from the type locality. This is the northernmost record of the genus and the first from the Bering Sea. According to observations from videos the species is locally common on pebble patches in low-relief silt and sand habitat at depths between 203 and 219 m. The sponge was often in association with several unknown species of hydroids, zoanthids, and the demosponge Stylocordyla borealis ( Loven, 1868) .
Etymology: The species name “kanuux” means heart in Unungan and the Bering Sea canyons are regarded as the heart of the Bering Sea by the Unungan people.
Discussion: There are 21 nominal species in Aaptos (Van Soest et al., 2005) . We compare A. kanuux with congeners from the North Pacific and the N-Atlantic; other known species are unlikely to be conspecific for zoogeographic reasons and therefore are not considered further. Spicule measurements of Aaptos species compared with A. kanuux sp. n. are given in Tab. 1. A . kanuux sp. n. is the first record of the genus from the Bering Sea, which might be a valid argument for specific distinctness based on zoogeographic reasons. Zoogeographically the closest congener reported is Aaptos niger Hoshino, 1981 from Japan. A. niger differs ecologically as it was recorded from the subtidal at a depth of only 15 m. Aaptos niger has only two categories of spicules while the new species has three. The choanosomal strongyloxeas are roughly half the length but twice the thickness of those of A. kanuux sp. n. Ectosomal spicules differ in geometry and sizes between the two species; ectosomal styles in A. niger are much longer than ectosomal tylostyles in A. kanuux sp. n. The only other record of the genus in the North Pacific is A. vannamei de Laubenfels, 1935 reported from California. As A. vannamei completely lacks strongyloxeas, it does not fit in the definition of, and consequently is not a member of Aaptos . A. vannamei has styles of an extremely wide size-range and styles to subtylostyles. A confident assignment of de Laubenfels’ species is not possible without a closer examination of the holotype. The type species Aaptos aaptos ( Schmidt, 1864) was originally described from the Mediterranean Sea, but there are also N-Atlantic records. According to redescriptions of the type species, A. aaptos has two size categories of strongyloxeas ( Kelly-Borges & Bergquist, 1994, Van Soest, 2002), in contrast with only one in A. kanuux sp. n., with the maximum dimensions somewhat smaller. Both species have subtylostyles as intermediate spicules, but subtylostyles in the type species are somewhat longer and considerably thicker. The most striking difference is that of ectosomal spicules: these are curved and frequently flexuous styles or subtylostyles in the type species but straight tylostyles with well rounded tyles in A. kanuux , the latter being much shorter but thicker than ectosomal spicules in A. aaptos .
Aaptos papillatus ( Keller, 1880) resembles our new species in growth form as it is hemispherical, but differs in hav- ing surface papillae. Differences in spiculation are the strongyloxeas in a larger size-range in A. papillatus ; notably, the width of spicules being over twice that observed in A. kanuux sp. n. The same is true for the dermal tylostyles, which may be up to 36 µm thick compared to only 8 µm in A. kanuux sp. n. The third category of spicules is composed of styles in A. papillatus and subtylostyles in A. kanuux sp. n.; the maximum length of these is 770 µm in the new species compared to 1478 µm in A. papillatus . A. papillatus is a Mediterranean species, with one record from the Atlantic coast of France. This record was first described as Polymastia glenei by Descatoire (1966) but was later synonymised with A. papillatus . Again, the distance between the European coasts, either Mediterranean or Atlantic, argue for specific distinctness. A. bergmanni de Laubenfels, 1950 is a shallow water species described from Bermuda. It is a rather large species, dark brown outside and yellow in the interior. It differs in having ectosomal styles which are much thinner than the tylostyles of A. kanuux sp. n. Furthermore, the strongyloxeas in A. bergmanni are shorter and the species lacks a third category of megascleres.
To summarize, A. kanuux sp. n. appears to be unique with its mustard yellow exterior, and it differs from all congeners in the assemblage of spicules occurring and/or in their dimensions. Most species of the genus occur in warm or temperate seas and in shallow water, A. kanuux sp. n. being an exception for it was found in rather deep water in a sub-polar region.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |