
Euprymna brenneri, Sanchez & Jolly & Reid & Sugimoto & Azama & Marlétaz & Simakov & Rokhsar, 2019
|
publication ID |
https://doi.org/ 10.1038/s42003-019-0661-6 |
|
DOI |
https://doi.org/10.5281/zenodo.3728602 |
|
persistent identifier |
https://treatment.plazi.org/id/03E00C24-FFEA-CE16-257E-0D8DFE71F8E0 |
|
treatment provided by |
Plazi |
|
scientific name |
Euprymna brenneri |
| status |
sp. nov. |
Euprymna brenneri sp. nov.
(LSID: urn:lsid:zoobank.org:pub:B2D2A34E-FB8C-4D45-824E-9530986C6D44 ; Figs. 1c View Fig right, 4–9; Table 1 View Table 1 ; Supplementary Fig. 1 View Fig ; Supplementary Tables 2 and 3, and Supplementary Data)
Material examined. HOLOTYPE 1 ♂, 14.9 mm ML, immature, Okinawa, Diamond Beach in Seragaki, 26.51N, 127.88E, 26 Apr. 2016, coll. J. Jolly & O. Simakov ( NSMT Mo 85885 GoogleMaps ). PARA- TYPES Okinawa, Seragaki, Diamond Beach , 26.51N, 127.88E, 26 Apr. 2016, coll. J. Jolly & O. Simakov: 1 ♂, 18.8 mm ML, ( NSMT Mo 85886); GoogleMaps 1 ♂, 9.0 mm ML, immature ( NSMT Mo 85887); GoogleMaps 1 ♂, 10.6 mm ML, immature ( NSMT Mo 85888); GoogleMaps 1 ♀, 19.5 mm ML, immature ( NSMT Mo 85889); GoogleMaps 1 ♀ 11.6 mm ML, immature ( NSMT Mo 85890). Okinawa, Miyagi Is , 26.38N, 127.99E, <2 m: GoogleMaps 1 ♂, 22.0 mm ML, mature, 18 Mar. 2016, coll. B. Grasse, J. Jolly, O. Simakov, S. Nyholm, K. Keunig, A. Masunaga, F. Marletaz ( NSMT Mo 85891, Hap 23, and GenBank accession number: LC417234 View Materials ); GoogleMaps 1 ♀, 9.1 mm ML, immature, 24 Feb. 2018, coll. G. Sanchez, J. Jolly, C. Sugimoto, K. Asada ( NSMT Mo 85892, Hap 12, and GenBank accession number: LC417223 View Materials ). Okinawa, Kume Is, 23.33N, 126.81E, GoogleMaps 1 ♀, 15.3 mm ML, immature, 12 Nov. 2018, coll. R. Minemizu ( NSMT Mo 85893). GoogleMaps
Notes: Only one male was reproductively mature, but due to damage to the arms and missing suckers this specimen was not selected as the holotype. No females were reproductively mature. All specimens with the exception of NMST Mo 85891 and NMST Mo 85892 were laboratory-reared from collected eggs.
Other material examined. Syntypes: Euprymna bursa , Hong Kong, ♀ (34.4 mm ML), ZMH 1393 View Materials , ZMH 1384 View Materials ♀ (~ 25 mm ML) .
Diagnosis. Small species, largest examined specimen (male, mature) 22.0 mm mantle length. Arm suckers biserial proximally and at distal tips, rest tetraserial. Males with enlarged suckers in dorsal and ventral rows of arms 3 and 4 (from ~5th to 8th sucker row from base on arms 3 and from ~7th to 9th sucker row from the base of arms 4). Hectocotylus without finger-like papillae basally. Females with enlarged suckers in ventral row of arms 3 (from ~8th to 13th sucker row from the arm base) and dorsal and ventral row of arms 4 (from ~10th sucker row counting from the basal-most row of suckers). Male enlarged suckers relatively larger than female arm suckers; suckers on ventral side of arm 3 larger than remaining enlarged suckers in both sexes. Enlarged suckers not in continuous rows, but interspersed with small suckers.
Description. Counts and indices for individual specimens are given in Supplementary Table 3.
Species relatively small: ML males examined 8.6–22.0 mm ML, females 8.5–19.5 mm ML. Mantle short, broad, domeshaped posteriorly ( Fig. 4a, b View Fig ); MWI males 63.6–98.8, females 62.1–94.1. Dorsal mantle joined to head by broad commissure, ventral mantle margin wide “m” shape ( Fig. 4b View Fig ). Fins oval; length approximately 50% ML; FIIa males 26.4–31.9, females 25.6–32.9; fin width about 30% ML, FWI males 23.9–34.9, females 21.6–45.8, attached dorso-laterally midway along mantle. Fins do not project anteriorly as far as anterior mantle margin ( Fig. 4a, b View Fig ).
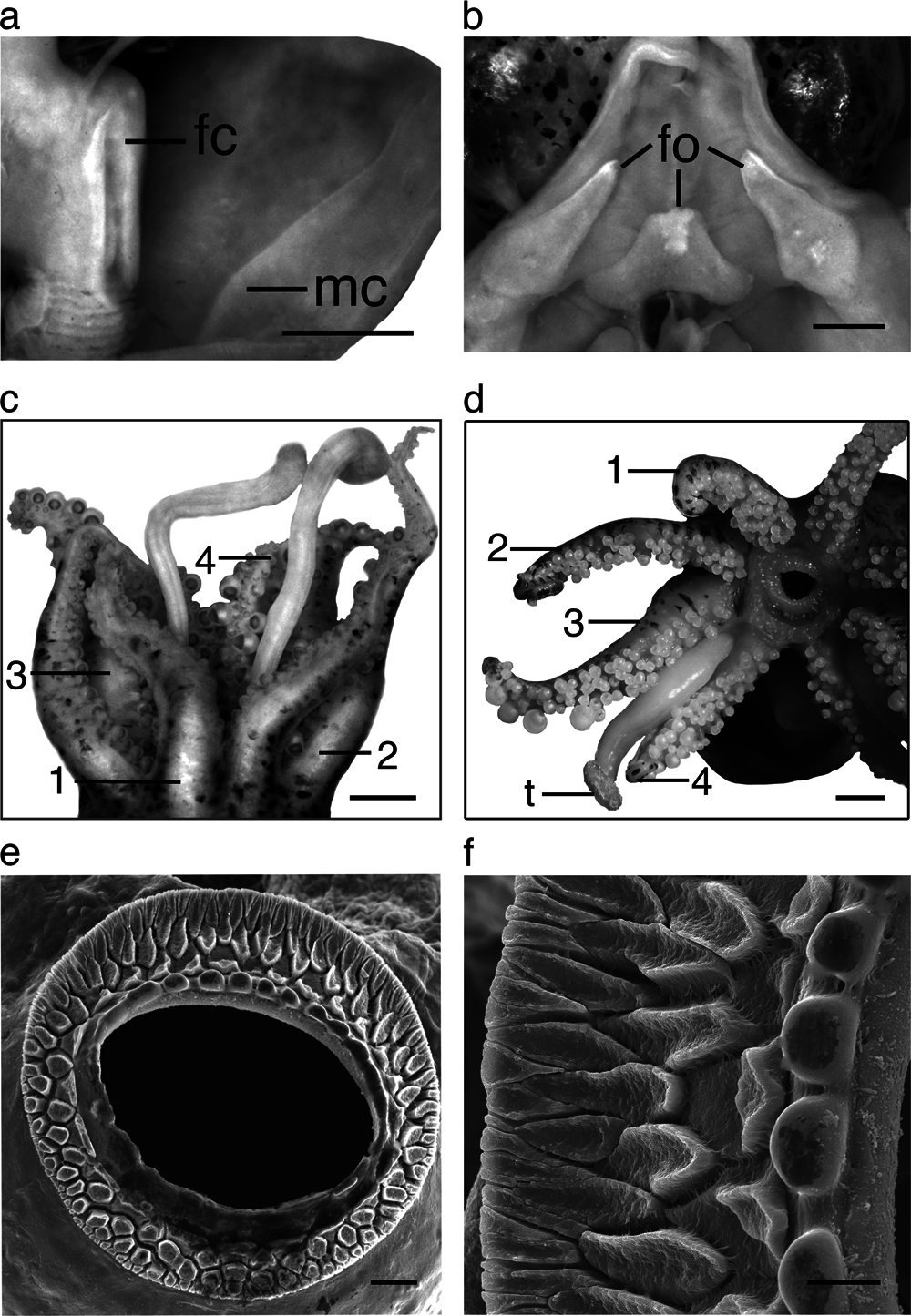
Funnel conical, projects anteriorly beyond ventral mantle margin ( Fig. 4b View Fig ). FuLI males 52.3–75.5, females 52.3–67.2; free for most of its length, FFuI males 27.3–47.2, females 33.3–41.2. Funnel-locking cartilage long, narrow, parallel-sided with shallow groove of uniform depth and thickened rim. Mantle cartilage a long straight ridge ( Fig. 5a View Fig ). Funnel valve a tiny flap. Funnel organ ( Fig. 5b View Fig ) dorsal element broad, inverted V-shape, with concentration of glandular tissue mid-anteriorly; ventral elements with acute anterior tips.
Head broader than long, HLI males 47.0–69.8, females 51.7–65.4; HWI males 66.0–96.5, females 62.6–91.8. Eyes large, EDI males 11.6–23.6, females 14.4–20.7; ventral eyelids free. Distinct, large photosensitive vesicle on latero-posterior surface of head, posterior and ventral to eyes.
Arms, broad basally, tapered distally; order 3> 1> 2 = 4 or 3> 2> 1 = 4 (Supplementary Table 3). Arm length index of longest arm in males (ALI3) 97.3, female (ALI3) 92.3; arm keels absent or indistinct. All arms connected by relatively shallow webs, protective membranes absent. Arm sucker pedicels without lappets. Arm suckers tetraserial, with some biserial suckers at base and at distal tips of arms; spherical on normal arms ( Fig. 5c, d View Fig ) (hectocotylus differs). Chitinous sucker rims: infundibulum with 3–4 rows of pavement-like processes ( Fig. 5e, f View Fig ), peripheral sucker rim processes wedge-shaped, rest irregular with slightly raised outer margins ( Fig. 5f View Fig ). Chitinous inner rim of normal arm suckers without teeth, slightly crenulated on one side ( Fig. 5f View Fig ). Suckers on modified region of hectocotylus with toothed margins ( Figs. 6 View Fig and 5a View Fig ).
Males ( Fig. 5c View Fig ) and females ( Fig. 5d View Fig ) with enlarged suckers on outer row(s) of arms 3 and 4. Enlarged suckers clearly discernible in both sexes, even those of the smallest sizes (male 8.6 mm ML, female 8.5 mm ML). Male enlarged suckers larger than female arm suckers (Supplementary Table 3). Sucker counts range from 60 to 112 on each arm; Arms 2 with a greater number of suckers than other arms in both sexes.
Males with arm suckers in the following variable arrangement from proximal to distal end of arm:
Right Arms 1 and Arms 2: 5 rows biserial suckers, rest tetraserial. None enlarged.
Arms 3: 5 rows biserial suckers, rest tetraserial, distal arm tip with ~6 rows biserial suckers. Dorsal rows distal to sucker rows ~5–7 with 4–9 enlarged suckers interspersed at intervals with regular-sized suckers. (Larger specimens with greater number of enlarged arm suckers.) Ventral rows with ~5 enlarged suckers interspersed at intervals with regular-sized suckers.
Arms 4: ( Fig. 5c View Fig ) 5 rows biserial suckers, rest tetraserial, distal tip of arm with ~3 rows biserial suckers. Dorsal rows distal to sucker rows ~7–9 with ~3–4 enlarged suckers interspersed at intervals with regular-sized suckers. Ventral rows after sucker rows ~7–9 with ~2–7 enlarged suckers, some alternating at intervals with regular-sized suckers.
Females with enlarged arm suckers in the following variable arrangement from proximal to distal end of arm:
Arms 1: 4–5 rows biserial suckers, rest tetraserial, distal 4 rows biserial. None enlarged.
Arms 2: 2–4 rows biserial suckers, rest tetraserial, distal 6 rows biserial. None enlarged.
Arms 3: 5 rows biserial suckers, rest tetraserial, distal tip of arm with ~8–10 rows biserial suckers. Dorsal rows without enlarged suckers. Ventral rows with ~8–13 rows of normal suckers proximally followed by ~4–5 enlarged suckers alternating at intervals (either large and small suckers alternate, or two large suckers alternate with 1–2 regular-sized suckers) toward distal half of arms.
Arms 4: ~8–9 rows biserial suckers, rest tetraserial, distal tip of arm with 4–5 rows biserial suckers. Dorsal rows with enlarged suckers in rows ~11–15. Ventral rows with enlarged suckers in rows ~10–14.
In both sexes the enlarged suckers on arms 3 are larger than those on arms 4 and those in the ventrolateral rows are larger than those on the dorsolateral rows. The enlarged suckers on arms 3 displace the regular-sized suckers laterally. (The arrangement of enlarged suckers varies considerably among the specimens examined and it would be useful when more material becomes available to map the arrangement of enlarged suckers in mature specimens of both sexes and over a range of specimen sizes to determine whether a clear pattern of regular and enlarged suckers can be discerned.).
Left dorsal arm of males hectocotylised: from base to distal end of arm, one single sucker, ~seven transverse rows of normal tetraserial suckers, remaining suckers with swollen pedicels, that form palisade arrangement ( Fig. 6 View Fig ), biserial with integument partially covering chitinous sucker rim in cap-like arrangement. No finger-like papillae at base of hectocotylised arm. Right dorsal arm of males with transverse rows of “normal” tetraserial suckers, with swollen pedicels.
Tentacles slender, stalks naked, semicircular in section. Club relatively short; ClLI males 18.6–33.0, females 17.9–32.9, recurved in preserved specimens, tapers to blunt end distally; suckerbearing face convex. Suckers ~0.04–~ 0.08 mm diameter in center of club; arranged in ~16–24 crowded oblique rows ( Fig. 7b View Fig ). Swimming keel on aboral side of carpus broad, extends posteriorly well beyond carpus. Club sucker dentition ( Fig. 7c, d View Fig ): inner ring without teeth; infundibulum with three rows of pavementlike processes; inner rows sub-rectangular, narrowing toward central opening; middle and outer rows ovoid; irregular, with strongly crenulated and pitted surface.
Well-developed light organ present overlying and associated with ink sac ( Fig. 7e View Fig ). Individual lobes rectangular bulb-like anteriorly, slightly enlarged; rounded posteriorly.
Gills with 24–27 lamellae per demibranch. Buccal membrane with six lappets; suckers absent.
Radula with seven transverse rows of teeth ( Fig. 7f View Fig ). Rhachidian teeth simple, without cusps, triangular, slightly concave laterally and ventrally. First lateral teeth similar in size and shape to rhachidian teeth with pointed cusps displaced laterally and directed toward midline of radula. Second and third laterals with elongate bases, longer, curved. Third laterals with scythe-like teeth, longer than second laterals.
Upper beak ( Fig. 7g View Fig ) with pointed rostrum, hood curved, high above crest posteriorly; jaw angle approximately 90°; lateral wall edge with slight indentation. Lower beak ( Fig. 7h View Fig ) with blunt protruding rostrum, rostral edge obtuse, curved, without distinct inner angle; hood notch absent, wings almost straight. Distinct dark pigmentation restricted to rostrum and hood of upper and lower beaks.
Gladius absent.
Spermatophores (fully developed only in NSMT Mo 85891) approximately ½ mantle length. Sperm reservoir contains coiled sperm cord ( Fig. 8a View Fig ). Cement body unipartite; cylindrical, approximately uniform width, connects to sperm reservoir via a broad duct ( Fig. 8a, b View Fig ). Oral end of ejaculatory apparatus with 3–4 simple coils. Male reproductive tract similar in structure to congeners ( Fig. 8c View Fig ). Spermatophoric gland with very large, bulbous terminal portion.
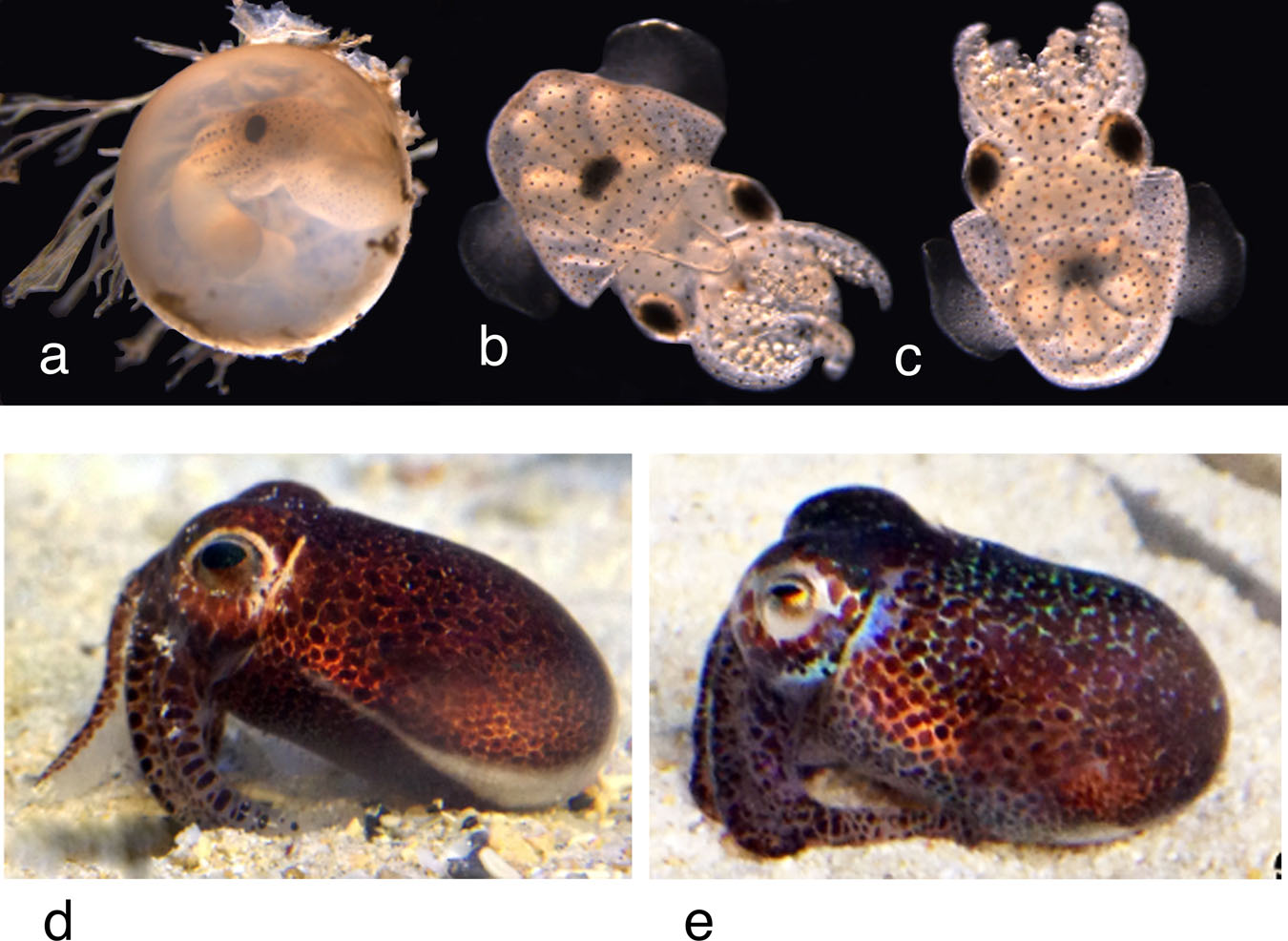
Female reproductive tract: Ovary occupies large proportion of posterior end of mantle cavity and opens via single thick-walled oviduct at anterior end on left side. Nidamental glands paired, broad, located ventral to ovary toward anterior end. Inverted brownish-colored U-shaped accessory nidamental glands located toward distal end of nidamental glands. Large sac-like bursa copulatrix on animals’ left side. Spawned eggs 4 mm diameter with jelly coat and sand, 3 mm without jelly coat. Eggs are laid in clusters of more than 100, rather than individually ( Fig. 1b View Fig , Type 3). Hatchlings 2 mm ML ( Fig. 9a–c View Fig , Table 1 View Table 1 ).
Color. Alcohol preserved specimens cream to maroon with large deep purple spots on dorsal and ventral head and mantle; spots larger and animal darker on dorsal surface ( Fig. 4a, b View Fig ); few scattered chromatophores on arms. Shiny bluish iridophores on head around eyes. Fins with large spots dorsally, close to junction with mantle, otherwise chromatophores absent from fins dorsally and ventrally. Club without pigment spots. Live adults rust brown with evenly scattered, relatively small pigment spots, darkest dorsally ( Fig. 1c View Fig Type 3; Fig. 9d, e View Fig ). Bright bluish iridophores around eyes, along anterio-dorsal rim of mantle, and underlie pigment spots on dorsal mantle ( Fig. 9e View Fig ). Arms banded with regular large spots and bars along their length ( Fig. 9d View Fig ) that can be seen even in embryos inside the eggs ( Fig. 9a View Fig ). Hatchlings translucent with evenly scattered chromatophores. Juveniles dark brown.
Habitat. Adults were found in sandy near-shore shallow waters, less than 2 m in depth, among corals and rocks. Eggs were found in rocky areas near coral reefs in depths of 8–18 meters.
Type locality. Japan ,
26.51N, 127.88E.
Okinawa,
Seragaki,
Diamond
Beach,
Distribution. Japan: Okinawa Prefecture Seragaki, Diamond Beach 26.51N, 127.88E; Oura Bay, 26.53N, 127.74E; Miyagi Island, 26.38N, 127.99E; Ishigaki Island, Oganzaki 24.44N, 124.07E; and Kume Island, Northern Hatenohama Beach (26.35°N, 126.86°E) ( Fig. 1 View Fig ). Taiwan *: off Penghu waters on the west-northern side of Sha-kang Fishing Harbor 23.60N, 119.62E. Depth range 2– 18 m. (*Based on COI obtained from a single immature specimen.)
Etymology. The species is named in honor of the pioneering geneticist and Nobel Laureate Dr Sydney Brenner, founding president of the Okinawa Institute of Science and Technology. We also propose the common name Brenner’ s bobtail in English and Buren ā -mimika in Japanese.
Remarks. The largest male (22 mm ML) (and the only fully mature specimen) examined had damaged arm tips and most suckers missing. However, the arrangement of enlarged suckers was clear from the sub-mature specimens.
Prior to this study fifteen species of Euprymna were recognized25, although three of these ( E. bursa Pfeffer, 1884 , E. pusilla Pfeffer, 1884 and E. schneehagenii Pfeffer, 1884 ) are considered doubtful species by Norman and Lu47. Among the nominal species of Euprymna , the species whose geographic range most closely encompasses the known range of E. brenneri includes E. berryi , which has been found in warm temperate coastal waters from China and Taiwan, south to Hong Kong and Japan 53, and E. morsei , which is sympatric with E. berryi over its range and also occurs as far south as the Philippines and Indonesia. Both, however, differ in morphology and in molecular traits ( Figs. 2 View Fig and 3 View Fig ) from Euprymna brenneri . These three taxa ( E. berryi , E. morsei and E. brenneri ) differ in their COI and transcriptome signatures. Both E. berryi and E. morsei have enlarged suckers in males on the second arm pair (on the ventral
margin in E. morsei and the dorsal and ventral margins in E. berryi ), while E. brenneri does not. Kubodera and Okutani32 described E. megaspadicea , found in deep waters (200 m) of Nago Bay off Okinawa. While no sequence data currently exists for E. megaspadicea , it is clearly morphologically distinct from E. brenneri . In E. megaspadicea the hectocotylised arm is longer than the opposing arm, and the hectocotylus contains a sharp lateral inward groove not seen in other Euprymna species.
As part of this study we examined the type specimens of E. bursa (ZMH RK 1384 and RK 1393, both females, approximately 25 mm ML and 34 mm ML respectively) from Hong Kong. Eurymna bursa differs from E. brenneri in that none of the arm suckers are enlarged (luckily these remain attached to the arms of the E. bursa types, enabling this comparison to be made). In addition, E. bursa has a greater number of arm suckers (102–128) and the median component of the funnel organ of E. bursa is spade-shaped, straight posteriorly, and not indented. (Whether E. bursa is a valid species awaits the examination of males from the type locality; here we verified that E. brenneri was not referable to this species—particularly important given their geographical proximity.)
Of the remaining Euprymna species (which now also includes E. parva ), the results of the COI analyses ( Fig. 2 View Fig ) indicate that E. brenneri (i.e., Ryukyu Type 3) belongs in a clade distinct from E. berryi ; E. hyllebergi ; E. morsei ; E. pardalota ; E. scolopes , E. tasmanica , E. albatrossae Voss 1963 , and Euprymna sp. Type 1. Transcriptomes separate E. brenneri from E. berryi , E. morsei , E. parva , E. scolopes , and E. tasmanica and Euprymna sp. Type 1 ( Fig. 3 View Fig ). These differences are also supported by morphological traits: no other Euprymna taxa are yet known to include females with enlarged suckers. In addition, male E. albatrossae ; E. berryi , E. megaspadicea , E. morsei , E. scolopes , E. stenodactyla Grant, 1833 , and E. tasmanica have enlarged suckers on the second arm pair of males, which is not the case for E. brenneri .
Euprymna brenneri does, however, have enlarged suckers on arms 3 and 4. Males and females have enlarged suckers on the ventral row of the third arm and the dorsal and ventral rows of the fourth arms, with no enlarged suckers on the first and second arms. This is the first time a female Euprymna has been identified with large suckers. Female members of this genus are notoriously difficult to identify based on morphology, so the discovery of this character is a valuable one. Of the other nominal species, the enlarged sucker arrangement in male E. brenneri is most similar to that of E. hoylei Adam, 1986 , but this species (in addition to all other nominal Euprymna with the exception of E. brenneri ) have 1–3 enlarged finger-like papillae on the proximal end of the hectocotylised arm. Euprymna hoylei , described from the Sulu Archipelago, like E. brenneri , has no enlarged suckers on the second arm, however, E. brenneri males possesses approximately eight very large suckers on the third arm compared to a smaller number described for E. hoylei (3–4). Female E. hoylei do not have enlarged suckers. In addition to the presence or absence of enlarged suckers on particular arms, the enlarged suckers in both sexes of E. brenneri are not located close to the base of the arms, as seems to be the case in other Euprymna species, but at some distance distal to the arm bases.







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Sepiolinae |
|
Genus |