
Coryphomys, Schaub, 1937
|
publication ID |
https://doi.org/ 10.1206/692.1 |
|
persistent identifier |
https://treatment.plazi.org/id/FA68571D-3114-FFBB-FF52-33D893F886A5 |
|
treatment provided by |
Tatiana |
|
scientific name |
Coryphomys |
| status |
|
Coryphomys View in CoL , sp. indet.
A number of specimens are referred with confidence to the genus Coryphomys , but due to lack of direct association with cheek teeth, cannot be determined to species. Nevertheless, this material adds significantly to our knowledge of the genus and warrants description under a generic banner.
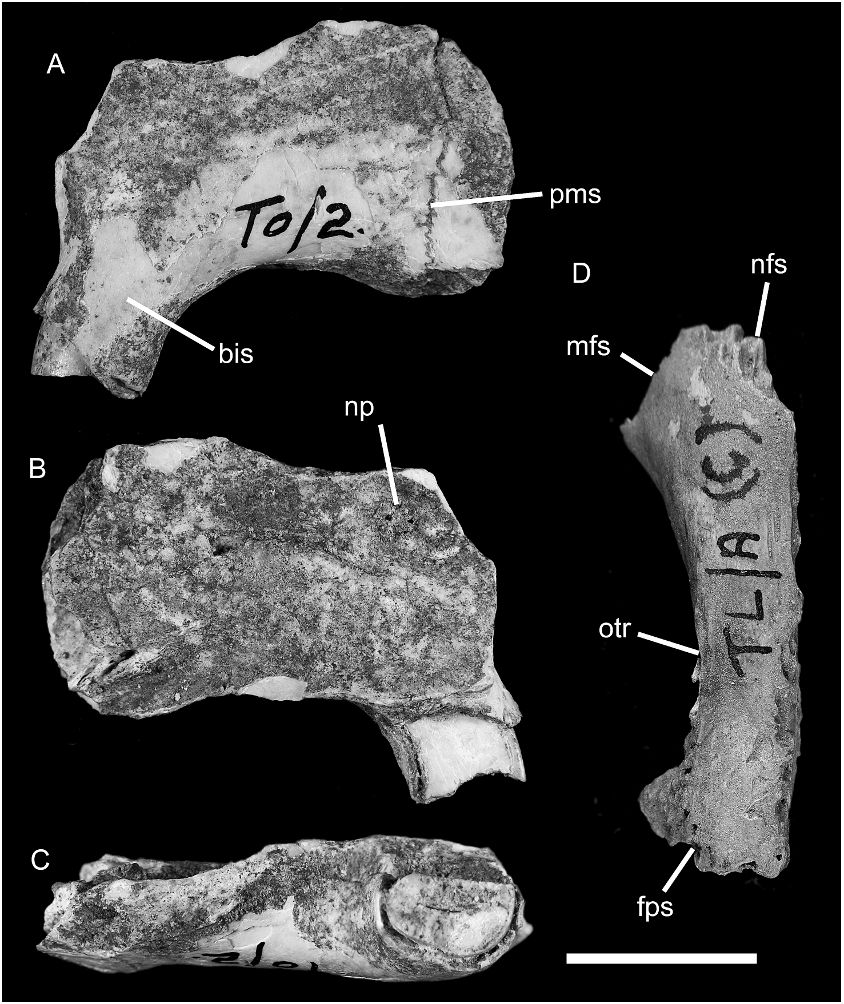
The most significant specimens are: AMF 68822: Uai Bobo 1 (TO/2), a partial left premaxilla; AMF 68760: Uai Bobo 1 (T0/3a), a fragmentary left zygomatic plate; AMF 68747: Uai Bobo 1 (T0/3a), a fragmentary right zygomatic plate; AMF 68864: Lie Siri (TL/A c), an isolated left frontal bone; ANWCP30: Matja Kuru 2 (MK2/D/31), an isolated right petrosal with minor abrasion; ANWCP31: Matja Kuru 1 (MK1/D/31B), an isolated and burnt left petrosal missing most of the pars cochlearis.
PREMAXILLA: A partial left premaxilla (AMF 68822; fig. 17A–C View Fig ) is referred on the basis of its simple D-shaped incisor morphology (described below), which parallels that seen in the lower incisor. The specimen also differs in both incisor and bony morphology from a directly associated premaxilla of a species of Mahoney’s Genus A. The specimen retains the broken stump of the incisor and is complete from the narial rim back to the maxillary suture; this dimension measures 20.44 mm. The palatal process is abraded such that the symphysis and details of the incisive foramen cannot be discerned. The dorsal suture between the narial process and the nasal bone is also lost.
The surviving portion, oriented with reference to the anterior surface of the incisor, suggests a relatively narrow rostrum that is not expanded lateral to the incisor gyrus (contra examples referred to Mahoney’s Genus A). The bony sheath of the incisor projects forward and downward from the palatal surface of the premaxilla, suggesting a weakly proodont incisor orientation (sensu Thomas, 1919).
UPPER INCISOR: This tooth in AMF 68822 has a simple D-shaped cross-section ( fig. 17C View Fig ). It measures 5.51 mm in depth and 3.55 mm in width, giving a depth to width ratio of 1.57. Pale orange enamel is restricted to the anterior half of the outer surface. The inner surface of the tooth is flat; the posterior surface is rounded.
ZYGOMATIC PLATE OF MAXILLA: Two fragmentary zygomatic plates (AMF 68760 and AMF 68747, not shown) are allocated to Coryphomys on the grounds of their close resemblance to this structure in a specimen referred to C. buehleri . In both the plate is tall and narrow, with a trapezoidal shape, and the dorsal margin displays the unusual feature of a strongly rugose and weakly overhanging posterior margin of the masseteric fossa. AMF 68760 preserves the full ventral root of the plate, which measures 10.2 mm in length, and a slightly damaged dorsal root with an indicated width of 6.05 mm. The height of the plate, measured from the inferior notch of the anterior infraorbital fissure to the highest point on the superior rim of the masseteric fossa, is 12.9 mm.
FRONTAL: An isolated left frontal bone (AMF 68864; fig. 17D View Fig ) is referred to Cor- yphomys. The specimen is virtually complete save for some damage to the ventral margin of the orbital lamina.
The dorsal surface of the frontal is longitudinally rather flat, but it shows a very slight midline doming anteriorly and a slight midline depression posteriorly. The orbitotemporal ridge is sharp posteriorly where it narrowly overhangs the orbitotemporal fossa. It fades anteriorly as it passes onto the rostral portion of the bone. The minimum interorbital width (midline suture to orbitotemporal ridge) is 5.5 mm, giving a full interorbital width of 11 mm. The anterior margin of the frontal is a simple V-shape with the apex facing anteriorly. This margin is comprised of two vertical, interdigitated sutures—the medial side is the nasofrontal suture, while the lateral suture is the maxillofrontal suture. The former suture is more finely interdigi- tating than the latter. The posterior margin of the frontal also bears a composite suture, the medial portion representing vertical, interdigitated contact with the parietal, and the lateral portion bearing a flattened facet for an overlying cranial lamina of the squamosal.
The orbital lamina of the frontal shows a relatively weak vertical fossa marking the broad fleshy origin of the orbital part of the temporal muscle. The ethmoid foramen is not present in the surviving part of the lamina and there is no sign of a dorsally placed foramen for the frontal diploic vein.
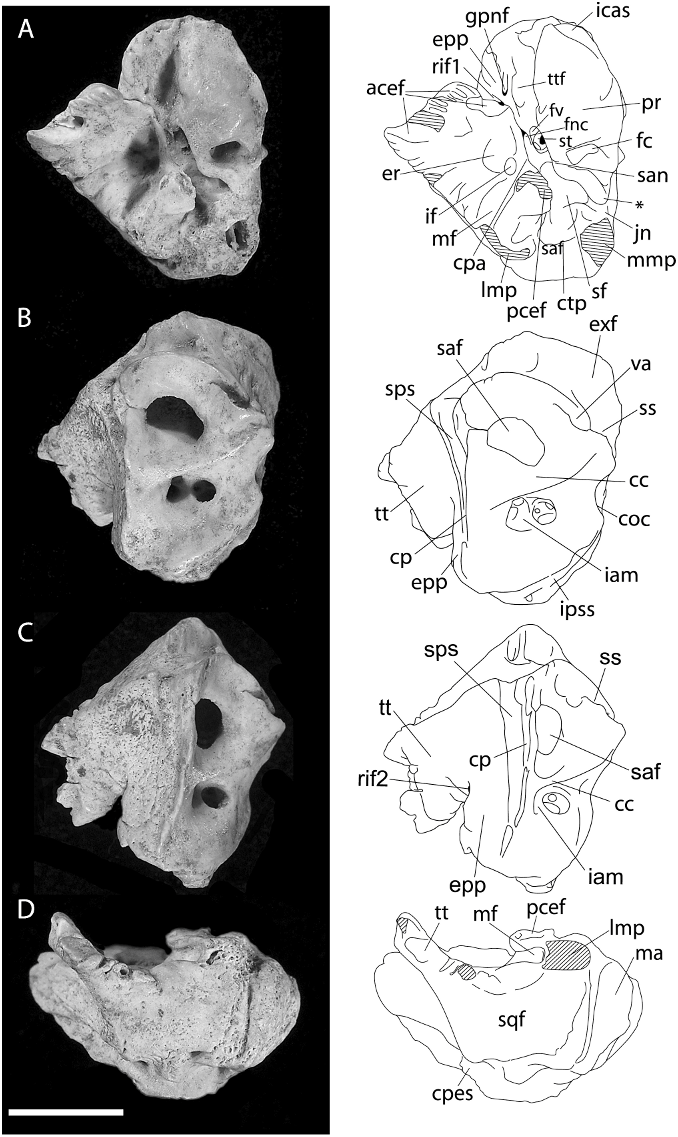
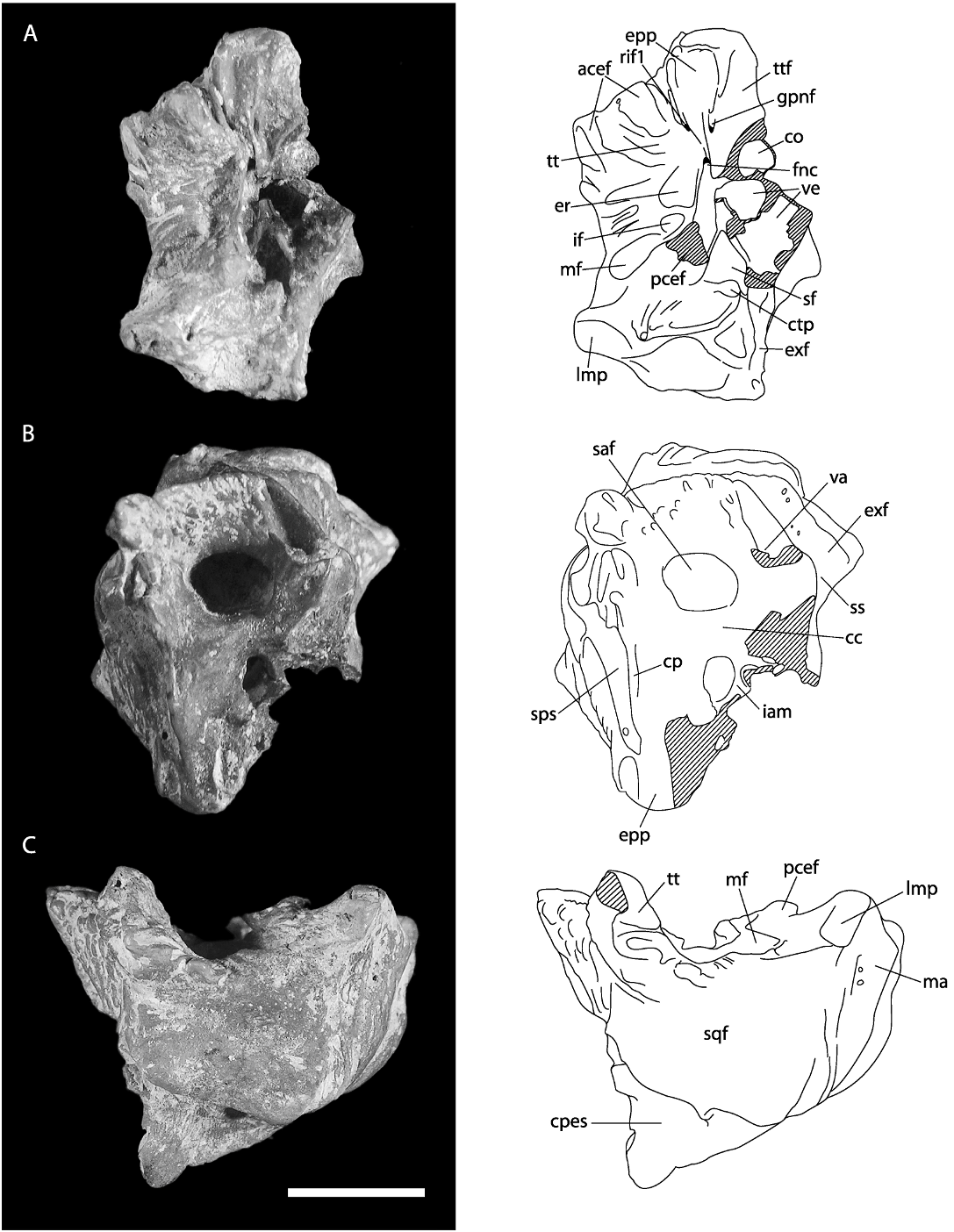
PETROSAL BONE ( figs. 18–19 View Fig View Fig ): The more complete of two specimens referred to Coryphomys ( ANWCP30 ; fig. 18A–D View Fig ) is a right petrosal with slight abrasion of the mastoid surface and the medial margin of the epitympanic wing. Important features of the tympanic surface ( fig. 18A View Fig ) are: (1) presence of a broad sulcus on the anterior pole of the pars cochlearis, presumably marking the passage of the internal carotid artery to the carotid canal; (2) absence of an obvious sulcus for the stapedial artery (the passage of a much reduced stapedial artery is possibly indicated by a small notch in the horizontal bony ridge that emanates from the ventral margin of the fenestra vestibuli and forms the medioventral lip of the stapedius fossa); (3) deep excavation of the tensor tympani fossa into the anterolateral surface of the promon- torium; (4) incomplete fusion of the tegmen tympani and the epitympanic process of the petrosal above the facial nerve canal; (5) presence of very small foramina within the zone of union of the tegmen tympani and epitympanic process of the petrosal, one on each of the tympanic and endocranial surfaces (both are distinct from the foramen for the greater petrosal nerve, which opens onto the tympanic surface of the epitympanic process of the petrosal); (6) robust nature of the tegmen tympani, supporting a broad area of fibrous attachment to the anterior crus of the ectotympanic (not preserved); (7) deep excavation of the stapedius fossa above a bony ridge that links the ventral margin of the fenestra vestibuli and the medial end of the caudal tympanic process; (8) presence of a shallow epitympanic recess with a distinct incudal fossa at its rear; (9) presence of an elongate, transversely oriented meatal fossa, presumably marking the limits of attachment of the pars flaccida of the tympanic membrane; (10) weak development of the caudal tympanic process behind the posterior tympanic sinus; and (11) medial end of the posterior tympanic sinus is partially enclosed by a thickened bony lamina that narrows the connection with the jugular notch.
The medial and endocranial surfaces ( fig. 18B–C View Fig ) show several features of note: (1) a broad, rugose surface marking the zone of contact with the exoccipital; (2) a broad sulcus for the sigmoid sinus, leading to a prominent jugular notch; and (3) a short sulcus, anterior of the vestibular aqueduct, presumably for the inferior petrosal sinus. The endocranial surface also presents two notable features: (1) a conspicuously large vestibular aqueduct (for the endolymphatic duct) passing into the body of the pars vestibularis of the petrosal behind and slightly above the subarcuate fossa; and (2) a broad, deeply impressed sulcus running anterior to the crista petrosa. Though the latter sulcus is located in an appropriate position to mark the passage of a prootic sinus, this vessel appears to be lost in later development of all placental mammals studied by embryological methods to date ( Wible, 1990; Wible and Hopson, 1995), with its drainage diverted into a capsuloparietal emissary vein (5 petrosquamous sinus of human anatomy; e.g., Diamond, 1992). In the Coryphomys petrosal a sulcus for the capsuloparietal emissary vein is visible in lateral view ( fig. 18D View Fig ), directed as in other murines toward a postglenoid foramen located between the petrosal and the squamosal bones. Given this observation, the additional sulcus in Coryphomys most likely marks the course of an enlarged superior petrosal sinus (connecting the cavernous sinus with the transverse sinus; Greene, 1968). Another feature of note seen in lateral view is a small, unidentified vascular channel that emerges out of the posterior tympanic sinus behind the lateral mastoid process.
The vascular impressions on ANWCP30 advertise an arterial arrangement characterized by strong reduction of the stapedial artery and its branches, and by a largely extracapsular course for the internal carotid artery. This is a derived arterial arrangement among muroid rodents but is one that has evidently originated on a number of separate occasions across several families (see Discussion). Much less comparative information is available on the pattern of endocranial venous drainage of muroids. However, our preliminary comparisons have failed to find a parallel morphology in any other murine taxon.
The second specimen (ANWCP31, fig. 19 View Fig ) is a burnt left petrosal missing most of the pars cochlearis. This specimen is noticeably larger than the more complete example, but it is otherwise very similar in morphology. Most notably, it displays the highly diagnostic feature of the broad endocranial venous sulcus anterior to the crista petrosa. One minor point of difference is the more prominent development of the lateral mastoid process, though we note that that this process is somewhat abraded in ANWCP30. The larger specimen is tentatively referred to C. buehleri and the smaller specimen to C. musseri .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.