
Chromaspirina guanabarensis, Maria, Tatiana Fabricio, Esteves, André Morgado, Smol, Nicole, Vanreusel, Ann & Decraemer, Wilfrida, 2009
|
publication ID |
https://doi.org/10.5281/zenodo.187502 |
|
DOI |
https://doi.org/10.5281/zenodo.6218807 |
|
persistent identifier |
https://treatment.plazi.org/id/F6288789-2164-FFD4-FF7F-16D427E4FBDD |
|
treatment provided by |
Plazi |
|
scientific name |
Chromaspirina guanabarensis |
| status |
sp. nov. |
Chromaspirina guanabarensis sp. n.
Figs 1 View FIGURE 1 and 2 View FIGURE 2
Type specimens: all in the Museu Nacional do Rio de Janeiro: holotype male ( MNRJ 305), two paratype males ( MNRJ 307), allotype female ( MNRJ 306), two paratype females ( MNRJ 308) and three paratype juveniles ( MNRJ 309).
Type locality: Bica Beach (22°49’37.9’’S and 43°11’15.9’’W), Guanabara Bay, Rio de Janeiro, Brazil, collected on 08/06/2000 by Dr. André Esteves. Sediment of very coarse to coarse sand with median grain size ranging from 2850µm to 570µm; intertidal zone.
Etymology: The species name refers to the type locality.
Measurements: Table 1.
-: not applicable or not seen.
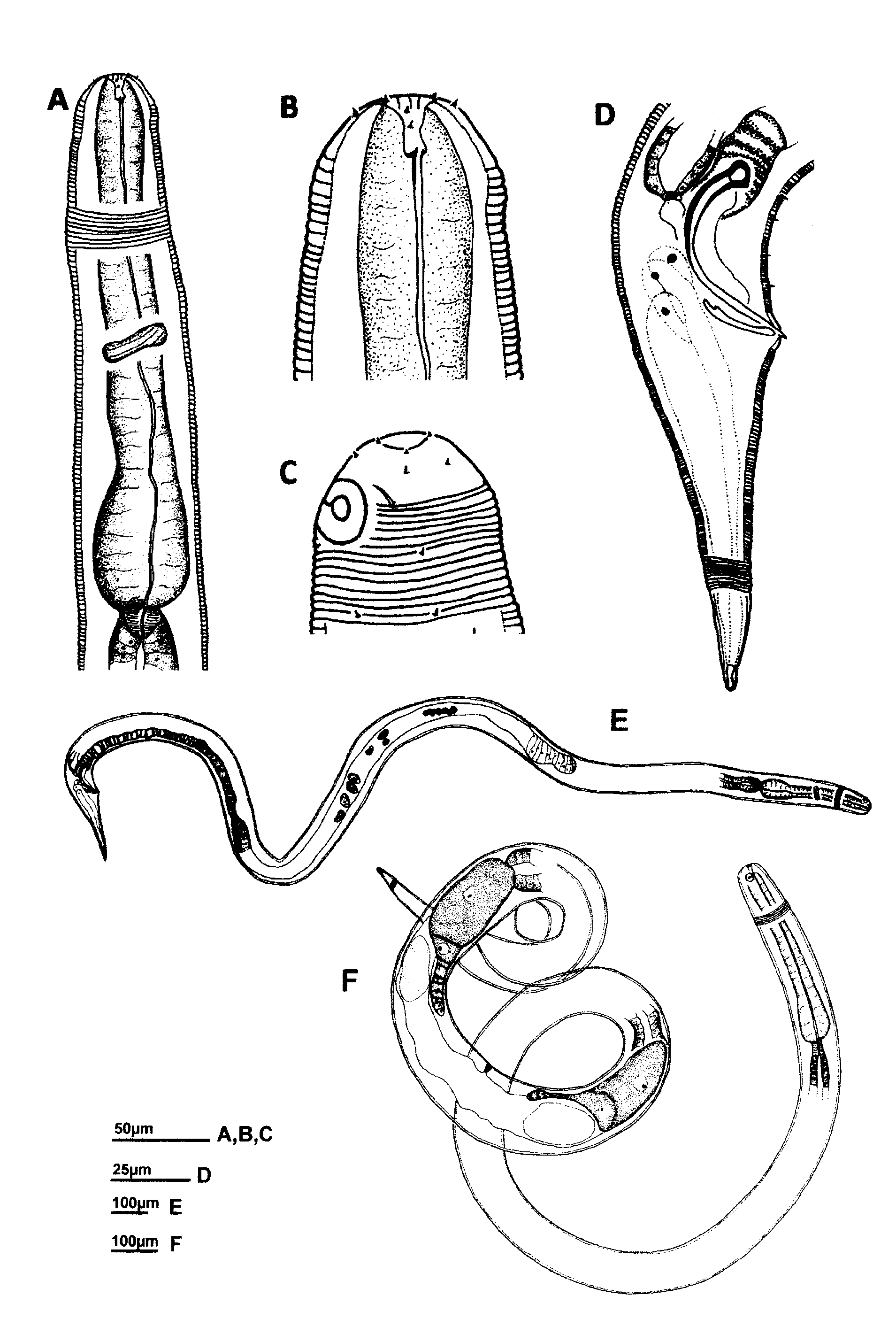
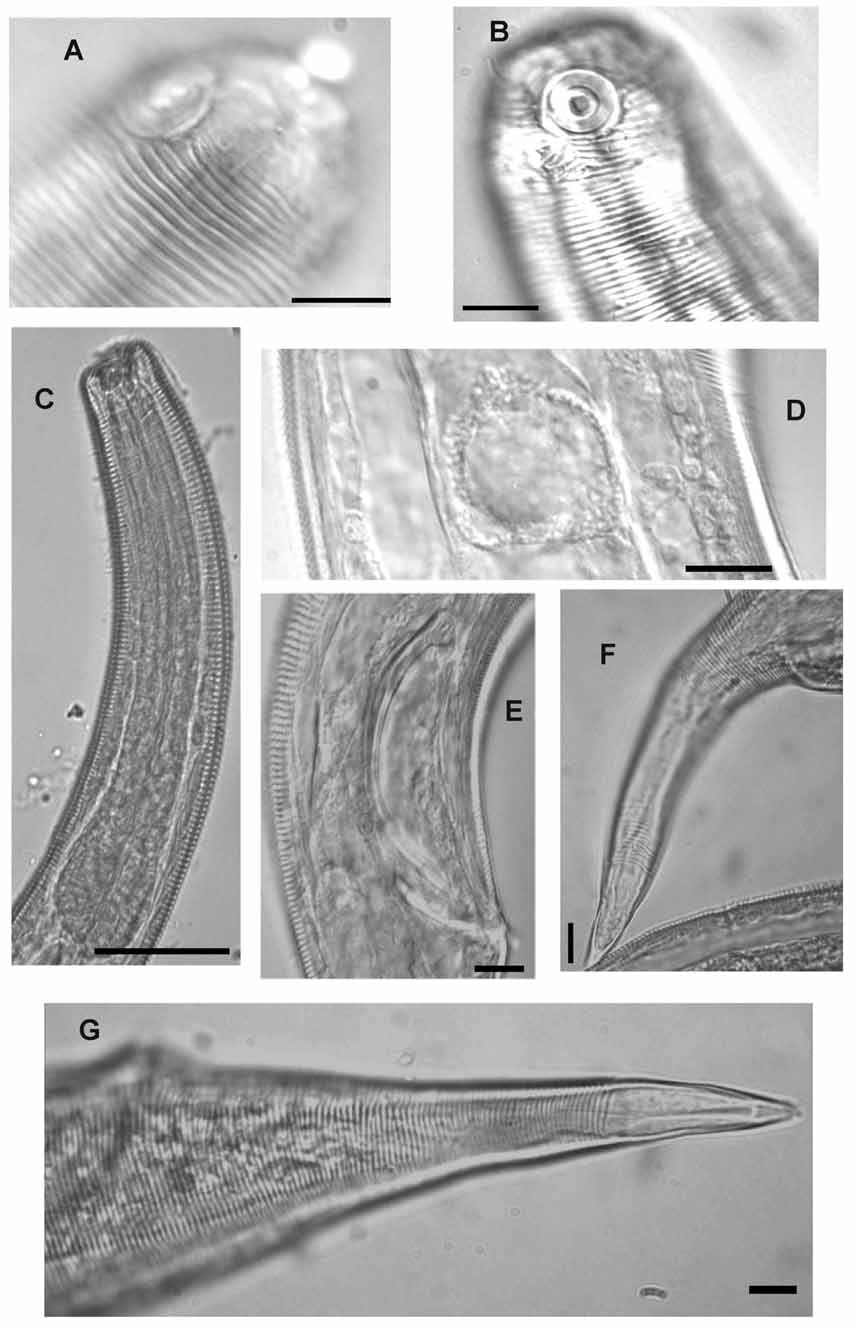
Male. Body long and relatively slender. Head blunt, rounded. Tail conical. Cuticle with fine transverse striations starting posterior to the anterior edge of amphidial fovea and without ornamentation. Minute somatic setae arranged in eight longitudinal rows extending from the pharyngeal region to tail region. Head region smooth, with six inner and six outer labial papillae, and four cephalic setae 4–5 µm long, located just anterior to the anteriormost striation of the body cuticle. Amphidial fovea a ventrally wound spiral, largely surrounded by striations; its diameter 29–39% of corresponding body diameter. Buccal cavity large with an acute dorsal tooth and two smaller ventrosublateral teeth. Pharynx anteriorly cylindrical, posteriorly widened to a pyriform bulb. Nerve ring located at about mid-pharynx length. Cardia triangular in cross section, surrounded by intestine. Tail conical with three caudal glands dorsally located, extending up to the cloaca and opening at the tail tip through a spinneret. Terminal part of tail smooth, 26–28% of the total tail length. Reproductive system monorchic with anterior outstretched testis on right side of the intestine. Testis with narrow short germinal zone. Sperm cells round (16–21µm diameter). Spicules equal, slender, ventrally curved with thick sclerotized lamina and hook-shaped capitulum; thin velum present. Gubernaculum slightly arcuate, trough-shaped, without apophysis. No obvious precloacal supplements present, but equally spaced short ventral setae which could be interpreted as supplements.
Female. Similar to male except for a cylindrical cardia, a shorter smooth tail portion (17–24% of tail length). Reproductive system didelphic-amphidelphic with reflexed ovaries; anterior ovary to the left of intestine, posterior ovary to the right of intestine. Anterior genital branch shorter than posterior. Vulva a simple transverse slit. One large egg (78–82µm long and 38–44µm wide) present in each uterus.
Juvenile. Only second stage juveniles observed; this stage is characterized by its small body size and small primordium, not clearly observed. External morphology similar to the adults.
Diagnosis. Chromaspirina guanabarensis sp. n. is characterized by its large size (> 2mm); the position of the cephalic setae at the posterior edge of the smooth head region, and the conical tail shape in both sexes. The non-striated part of the tail is shorter in females than in males. Males are characterized by spicules shaped with hooked sclerotized capitula and well sclerotized lamina, together with absence of precloacal supplements but presence of short setiform precloacal sensilla.
Differential diagnosis. Chromaspirina guanabarensis sp. n. most closely resembles C. okemwai Muthumbi, Verschelde & Vincx, 1995 in having similar dorsal tooth and spicule shape, but differs in having a more evenly tapering tail and more pronounced labial papillae ( C. okemwai has a tail with a wide anterior part narrowing posteriorly and minute labial sensilla only visible with SEM).
Discussion. The species included in the genus Chromaspirina have changed frequently. We consider the genus to have twenty valid species (including the new species). There are two species inquirendae, eight species transferred back to their original genus, and two species transferred to another genus.
C. robusta Wieser, 1954 and C. paucispira Stekhoven, 1950 , known from two juveniles and one female respectively, were regarded as species inquirendae by Wieser & Hopper (1967). The genus Bolbolaimus was synonymized with Chromaspirina by Luc & De Conninck (1959) but re-established as a valid genus by Jensen (1978), based mainly on the female reproductive system with outstretched ovaries, stoma and pharynx structure and cuticle ornamentation. Jensen re-established C. denticulata (Gerlach, 1953) , C. pellucida ( Cobb, 1920) and C. punctata ( Cobb, 1920) within Bolbolaimus when rejecting synonymy of this genus with Chromaspirina ; and regarded two other species C. amabilis (De Man, 1922) and C. cobbi ( Chitwood, 1938) as species inqueridae of Bolbolaimus .
C. dimorpha ( Hopper, 1961) , C. inflexa (Wieser, 1951) and C. rabosa ( Gerlach, 1956) have been transferred back to their original genus Desmodora mainly because of the amphidial fovea not being surrounded by striations of the body cuticle ( Wieser & Hopper, 1967). Among the Chromaspirina species, the amphideal fovea is partially or totally surrounded by striations, e. g. C. pellita and C. parma , respectively. However, one exception occurs in the genus: C. dubia possesses striations restricted to the posterior quarter of the body, but other characteristics, such as a large dorsal tooth and a pyriform bulb support its presence within the genus Chromaspirina .
C. longisetosa Jensen, 1985 was transferred to the genus Bolbolaimus by Muthumbi et al. (1995) and C. spinulosa Wieser, 1959 was transferred to the genus Metadesmodora by Gerlach (1963), but Blome (1982) proposed the new combination Echinodesmodora spinulosa when he erected the genus Echinodesmodora .
C. inaurita is added to the list of valid Chromaspirina species; however it was not recorded by Muthumbi et al. (1995). C. inaurita possesses several characters (e.g. large stoma, pyriform pharyngeal bulb) indicating affinities with Chromaspirina , although it also shows some characteristics of the genus Spirinia , e.g. a small dorsal tooth. Within the genus Chromaspirina the dorsal tooth may be variable in size, so this character is insufficient to place the species outside Chromaspirina .
C. modesta is regarded as valid because the work of Bussau (1993) conforms to the code for zoological nomenclature is a valid publication, and has been recognised world-wide as a reference among the scientific community of marine nematodes researchers.
C. pontica and C. inglisi differ from each other only by the presence of fine hairs over the body in the latter. Warwick (1970) suggested that the hair-like ornamentations may be either easily overlooked or considered as epigrowths of unicellular cyanophyceous algae, and if so the species are the same. Muthumbi et al. (1995) describing C. okemwai also found some fine hair-like structures attached to the cuticle of their specimens, but when the organisms were sonicated and viewed by SEM, the tail region of some animals appeared devoid of the “hairs”. The hair-like structures were also present on pre-moult phase of the newly made cuticle ( Muthumbi et al., 1995). Therefore, the origin of these structures remains doubtful. We regard C. pontica and C. inglisi as valid species, but consider that the hypothesis of C. inglisi being a junior synonym of C. pontica should not be completely disregarded though further investigation is needed.
Four species in the genus have a smaller dorsal tooth when compared with the remaining 16 species. This emphasises the close relationship of this genus with Spirinia as suggested by Wieser & Hopper (1967). Further investigation is needed into the relationships Spirinia with Chromaspirina . Chromaspirina guanabarensis sp. n. belongs to the group of species characterized by a poorly developed dorsal tooth. Within this group, the new species is easily discriminated by the absence of sexual dimorphism in the amphidial fovea, distinct labial sensilla and tail shape.
| MNRJ |
Museu Nacional/Universidade Federal de Rio de Janeiro |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Chromaspirina guanabarensis
| Maria, Tatiana Fabricio, Esteves, André Morgado, Smol, Nicole, Vanreusel, Ann & Decraemer, Wilfrida 2009 |
C. longisetosa
| Jensen 1985 |
C. dimorpha (
| Hopper 1961 |
C. spinulosa
| Wieser 1959 |
C. rabosa (
| Gerlach 1956 |
C. robusta
| Wieser 1954 |
C. denticulata
| Gerlach 1953 |
C. inflexa
| Wieser 1951 |
C. paucispira
| Stekhoven 1950 |
C. cobbi (
| Chitwood 1938 |
C. amabilis
| De Man 1922 |
C. pellucida (
| Cobb 1920 |
C. punctata (
| Cobb 1920 |