
Dasya patentissima Pena-Martín, M.B. Crespo & Gómez Garreta, 2014
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.184.5.2 |
|
persistent identifier |
https://treatment.plazi.org/id/F5171371-FFF9-B476-FF18-C56EFA8DFC22 |
|
treatment provided by |
Felipe |
|
scientific name |
Dasya patentissima Pena-Martín, M.B. Crespo & Gómez Garreta |
| status |
sp. nov. |
Dasya patentissima Pena-Martín, M.B. Crespo & Gómez Garreta , spec. nov.
Species notabilis ab affinibus Dasya ocellata et Dasya baillouviana facile distinguitur ramulis brevioribus perpendicularibusque, patentissimis, quae in utrisque longioribus et erecto-patentibus, et praeter has in posteriore estatura valde minore et cystocarpis sessilibus sunt. Hi habito , ramulis et cystocarpis characteres in ceteris Dasyis conjuncte non invenitur et speciei novae fortiter fundant.
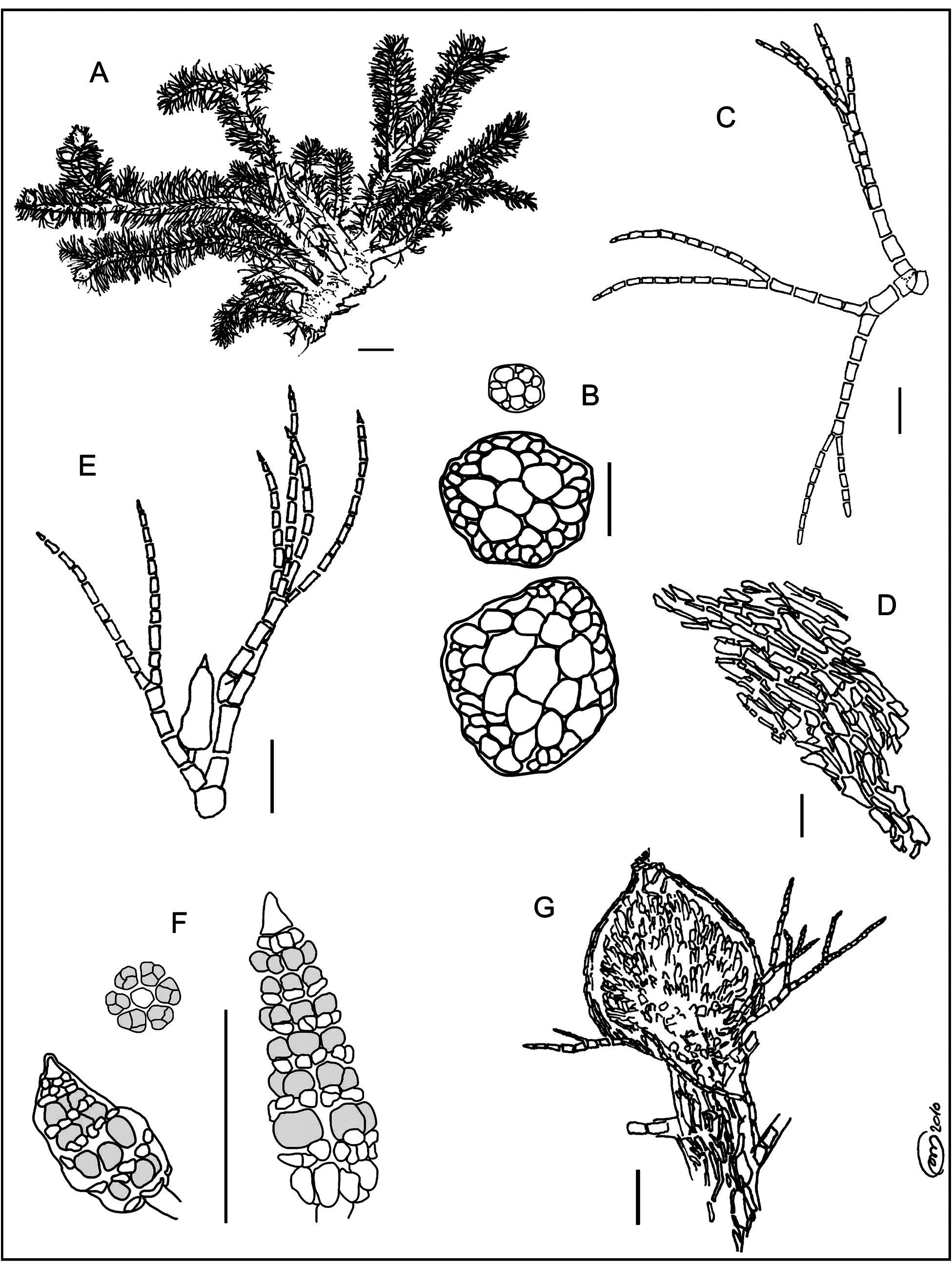
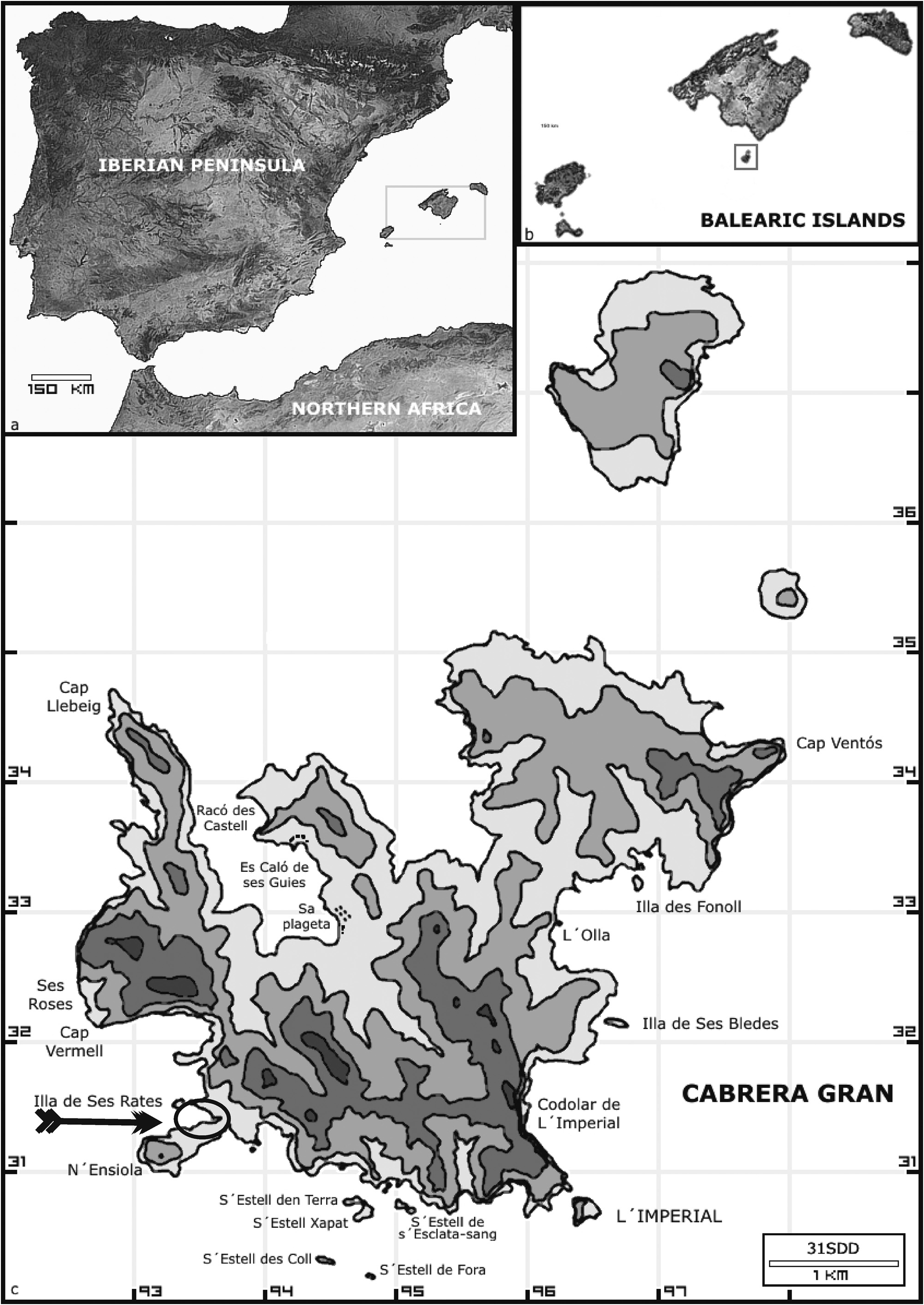
Type:— SPAIN. Balearic Islands: Cabrera Gran, Faro d’Ensiola , 39º 07’ 54” N, 02º 55’ 35” E (UTM: 31SDD9331), – 1 m a.s.l., 3 June 2005, M. Martínez-Azorín s.n. (holotype: ABH-Algae! num. 649) ( Figs. 1–15 View FIGURE 1 View FIGURES 2–8 View FIGURES 9–15 ) GoogleMaps .
Description:—Specimen c. 1 cm high, dark red and rigid texture ( Figs. 1A View FIGURE 1 and 2 View FIGURES 2–8 ); discoid rhizoidal holdfast 2 mm in diameter, bearing rhizoidal branches 400–900 μm long and 30–50 μm wide; 4 erect axes 200–475 µm wide, scarce and irregularly branched (1–2 times), and covered all over by patent uniseriate branches, deciduous near the base. The 5 periaxial cells of polysiphonous axes are clearly distinguishable in cross section near the apex ( Figs. 1B View FIGURE 1 and 3 View FIGURES 2–8 ), whereas near the base they are intermingled with cortical cells of similar size, this resulting in a cross section of polysiphonous axes apparently having 6–8 cells ( Figs. 1B View FIGURE 1 and 4 View FIGURES 2–8 ); axial and periaxial cells 60–100 µm long and 40–70 µm wide; polysiphonous axes corticated throughout, with elongated cortical cells (1–2 layers) with variable size (20–120 µm × 10–40 µm) ( Figs. 1D View FIGURE 1 and 5 View FIGURES 2–8 ). Pseudolaterals ( Fig. 6 View FIGURES 2–8 ) 400–800(1200) μm long, straight, and helicoidally ranged on polysiphonous axes, one per segment, with an angle of 80–90º; 3–4(5) dichotomies, according to the series: 1–2; 1–3; (2)3–4(5–8); (2)3–4(5–6)–end; 3–5(6)–end ( Fig. 1C View FIGURE 1 ); branchlets up to (8)11–15 cells long, closer to the apex; dichotomies forming 30–70º angles, except the first one (up to 90º) ( Fig. 1C View FIGURE 1 ); basal cell 50–70 µm long and 40–70 µm wide; suprabasal cell 60–75 μm long and 40–60 µm wide; fifth apical cell (numbering cells from the apex to the base of each pseudolateral) 25–40(100) μm long and 15–25 µm wide; apical cell 10–20(28) µm wide, but variable length, and rounded apex; at the apical zone of the thallus, apical cells closer (5–10 µm wide) and conical ( Fig. 7 View FIGURES 2–8 ). Some branches modified into rhizoidal filaments ( Fig. 8 View FIGURES 2–8 ). Numerous adventitious branches resembling pseudolaterals, and offering a whorled appearance ( Fig. 6 View FIGURES 2–8 ). Both adventitious branches and pseudolaterals are straight throughout, except near the apex, where they are slightly curved. Stichidia and cystocarps are found in the same individual ( Fig. 9 View FIGURES 9–15 ), though not spermatangial branches. Stichidia arranged in the apical part of the thallus, on first, second and/or third dichotomies of pseudolaterals, 1–2 per pseudolateral ( Figs. 1E View FIGURE 1 and 10 View FIGURES 9–15 ); 125–325 μm long and 45–90 μm wide, shape ovate-lanceolate ( Figs. 1F View FIGURE 1 and 11 View FIGURES 9–15 ); borne on 1(2)-celled pedicels, cells 20 µm long; conical apical cell ( Figs. 1F View FIGURE 1 and 12 View FIGURES 9–15 ) similar to the apical cell of the apex of pseudolaterals; each stichidium with 6–9 segments, 6 periaxial cells and 6 tetrasporangia per segment ( Figs. 1F View FIGURE 1 and 13 View FIGURES 9–15 ), 20–25 µm wide and partially protected by cover cells ( Fig. 1F View FIGURE 1 and 12 View FIGURES 9–15 ) which may be undivided or transversely or obliquely divided, and with irregular shape, 5–25 µm long and 5–25 µm wide, 10–16 per segment. Procarps not found. Cystocarps sessile, placed near the apex of the polysiphonous branches ( Fig. 14 View FIGURES 9–15 ). Pericarps urceolate, 350–500 μm long and 320–450 μm wide (excluding carpostome) ( Fig. 1G View FIGURE 1 and 15 View FIGURES 9–15 ); cells arranged in longitudinal rows or so, with various sizes and shapes, although generally elongated, 15–45 μm long and 10–25 wide. Carpostome scarcely prominent, 45–100 µm long and 50–70 µm wide. Gonimoblast with elongated carpospores (40–50 µm × 10–20 µm) ( Fig. 15 View FIGURES 9–15 ).
Etymology:—Named after the remarkable branching pattern, with patent branchlets, which is not found in any other Mediterranean species of the genus (patentissimus, -a, -um = very widely spreading, being arranged at an angle of 90º with respect to the main axis) ( Fig. 6 View FIGURES 2–8 ).
Habitat and distribution:— Dasya patentissima was found epilithic, growing in shallow depths (ca. 1 m deep), on rocky and protected shores, north of Punta d’Ensiola, southwest of Cabrera Gran (Balearic Islands) ( Fig. 16 View FIGURE 16 ). It occurred in sciophilous sites, under other larger algae, such as Halopteris scoparia ( Linnaeus 1753: 1165) Sauvageau (1904: 349) (≡ Conferva scoparia L.) ( Phaeophyceae), Halopteris filicina ( Grateloup 1806: unnumbered page) Kützing (1843: 293) (≡ Ceramium filicinum Gratel. ) ( Phaeophyceae) and Corallina elongata Ellis & Solander (1786: 119) (Florideophyceae) . It was found living together with two other congeners, Dasya ocellata and D. hutchinsiae . To date, D. patentissima appears to be limited to the type locality, where it probably is not very abundant, since later field work in Cabrera Gran did not yield new findings of the species.
Taxonomic relationships:— Dasya patentissima shows distinctive characters, which are not found in any of the known species of the genus, neither from the Mediterranean basin nor worldwide. The combination of smaller size, patent arrangement and short length of pseudolaterals, the apical location of cystocarps, and spatial branching with very wide dichotomies is unique and therefore diagnostic for the new species, all this allowing easy recognition. In particular, its morphologically closest Mediterranean relatives D. ocellata and D. hutchinsiae exhibit quite different features ( Table 1; Fig. 17–26 View FIGURES 17–26 ). It is also remarkable that D. patentissima produces six tetrasporangia per segment in the stichidia, instead of four or five as in both other congeners, and carpostome size is also smaller in the new species (length: 45–100 µm; width: 10–25 µm). Similarly, D. patentissima differs from D. ocellata by the presence of rhizoidal filaments, the smaller stichidia (125–325 × 45–90 µm), the contiguous (not separated) cover cells, and the smaller pericaps (350–500 × 320–450 µm) ( Table 1). From D. hutchinsiae , it is recognisable by the presence of adventitious branches, the complete cortication of thalli, the periaxial cells indistinguishable at the base, the narrower (15–25 µm) fifth apical cell of ramuli, the apical position of cystocarps, and the carpospores elongated in outline ( Table 1).
Although the type specimen of Dasya patentissima might be regarded as an atypical individual of D. ocellata or D. hutchinsiae , the fact that it is reproductive, all the morphological characters are constant through the thallus, and no aberrant specimens of both above cited species are known, allows us to rule out this possibility. Furthermore, the eventual inclusion of this specimen within the variation range of any of those two species would create a great distortion of the current taxonomic circumscription of both taxa (Pena-Martín 2011). All those reasons justify in our opinion the description of a new species.
Nonetheless, new collections of D. patentissima are necessary to undertake genetic analyses that will help to establish its taxonomic position and phylogenetic relationships. Two collecting attempts during the last years to find new localities of D. patentissima were unsuccessful to date. In the meantime, and due to the small amount of available original material, no DNA extraction was made to preserve the holotype.
So far, the available molecular papers that include Dasya species are focused on phylogenetic relationships within Florideophyceae, Ceramiales or some families close to Dasyaceae (cfr. Saunders & Bailey 1997; Jong et al. 1998; Choi et al. 2002, 2008; Kapraun & Dunwoody 2002), the sequences of Dasya they refer being only partial. Recently, Yamagishi et al. (2014) have reported a phylogenetic study on Dasya species from Japan, based mostly on the same data taken from GeneBank. Therefore, to date no comprehensive molecular studies are found allowing a proper discussion on phylogenetic relationships within the genus Dasya . Nonetheless, a preliminary phylogeny inferred from rbcL sequences (cpDNA) and morphological data including the west Mediterranean taxa of Dasya is underway.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |