
Chlamydoselachus africana, Ebert, David A. & Compagno, Leonard J. V., 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.189264 |
|
DOI |
https://doi.org/10.5281/zenodo.6223518 |
|
persistent identifier |
https://treatment.plazi.org/id/F41A87DE-FFDE-250C-C2C5-DDFEFAE731C5 |
|
treatment provided by |
Plazi |
|
scientific name |
Chlamydoselachus africana |
| status |
sp. nov. |
Chlamydoselachus africana View in CoL sp. nov.
Southern Frilled Shark
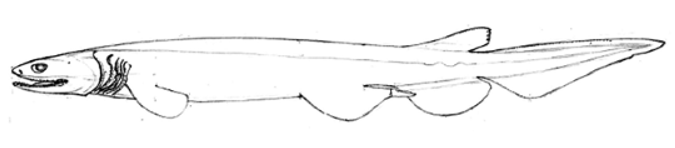
( Figures 1 View FIGURE 1 , 2 View FIGURE 2 , Table 1)
Chlamydoselachus anguineus: Smith 1951 View in CoL , 87; Smith 1965, 511, Fig. 3 View FIGURE 3 b; Smith, 1967: 105, pl. 19–23; Trunov, 1968, 137, fig. 3; Bass et al., 1975, 16, text fig. 9, pl. 6; Domanevskiy, 1975: 1117; Timokhin, 1980, 125; Allue et al. 1984, 124; Compagno 1984, 14, ill.; Bass 1986, 47, Fig. 3.1; Lloris, 1986, 87, figs. 17–18; Compagno et al., 1989, 19, pl. 2; Ebert 1990, 30, Fig. 3.1, Tab. 2.1; Compagno et al. 1991, 51, Fig. 3 View FIGURE 3 a (distribution map).
Chlamydoselachus View in CoL "sp. A" Ebert, 1990, 217, Fig. 3.12.
Chlamydoselachus "sp. A" Compagno et al., 2005, 65, Plate 1
Holotype. SAM 31028, 1170 mm TL, immature female, off the Cunene River, Namibia, 19° 59'S, 11° 48'E, 409 m deep, trawled by RV Benguela.
Paratypes. 4 specimens. RUSI 2423, 915 mm TL mature male, off Walvis Bay, Namibia; RUSI 2424, 1010 mm TL, immature female, off Walvis Bay, Namibia; SAM 36076, 916 mm TL, mature male, off Lüderitz, Namibia, 26°38'S, 15°10'E, 425 m deep, 19 February 1988; USNM 203466, 990 mm TL, mature male, Bahia Farta, Angola, 12°36'S, 13° 12'E, 5 April 1968.
Diagnosis. An external morphologically similar looking, smaller-sized Chlamydoselachus (to at least 1170 mm TL) with a proportionally long head length 17.3–17.9% TL, prepectoral length 15.1–17.7% TL, predorsal fin length 62.6–64.5% TL, head height 6.2–8.6% TL and width 6.6–8.9% TL, broad interorbital width (5.7–6.4% TL) and internarial width (3.9–4.8% TL), mouth broad (6.0–8.1% TL), gill openings long, analcaudal fin space (0.2–1.3% TL), pectoral fin length (8.8–9.6% TL), pelvic fin length (11.6–15.4% TL), mean number of upper tooth rows (x = 28.8), pectoral mesopterygial radial counts (7), metapterygial radial counts (8), anal fin radials (30), total vertebral counts (147), MP vertebral count (18), DP count (76), DC count (52), MP/DP transition just posterior to the pelvic fins, spiral intestinal valve counts 26–28.
Description. Proportional measurements expressed as a percentage of the total length are given for the holotype followed by the four paratypes in parentheses ( Table 1). Skeletal description and clasper morphology based on adult male paratype (SAM 36076). Body long, slender, eel-like, compressed behind the pelvic fins. Pectoral-pelvic space is moderately long with a noticeable difference between males 23.0–24.1% and females 26.8 (24.6–27.7)% of total length. Head broad, flattened, wider than high, slightly convex; head length 17.3 (17.4–17.9)% TL. Preoral snout length 0.1 times mouth width. Snout tip broadly rounded. Nostrils lateral, width 5.9 in internarial width, 2.2 in eye length, and 0.2 in third gill opening. Eyes large, rounded, length approximately 10.1 times in head length. Spiracle present or absent and when present about 13.3 times in eye length and located about 3.3 eye lengths behind posterior margin of eye. Distance from snout tip to sixth gill opening 1.5 times in pectoral-pelvic space. Height of gill openings descending in length, with the sixth being about 1.4 times height of first; first gill opening extends across throat; width of third gill opening about 2.4 times in head length and 4.3 times eye length. Mouth broadly rounded, large, distensible, length is about 0.9 times in mouth width; the width is about 2.7 times in head length, and 1.0 of head width at mouth corners.
Teeth are similar in both upper and lower jaws. Each tooth has three long, slender, smooth-edged cusps. Between each cusp is a small pointed cusplet. Upper medial teeth paired, form similar to anterolateral teeth, but noticeably reduced. Lower jaw with a single medial tooth that is undifferentiated from the anterolaterals. The teeth on upper and lower jaws are curved inwards and set on a broad base that projects behind and interlocks with the tooth base posterior to it. Tooth count for holotype is 14-2-14 / 13-1-13 ( Tables 1 & 2). There was no sexual dimorphism observed between the teeth of male and female sharks.
Lateral trunk denticles lanceolate, single cusped, with flattened bases. Crown slightly projected above the body with four longitudinal ridges extending from the base to the cusp. Denticle crowns widely spaced.
Pectoral fins are broad, rounded and low on body. Both anterior and posterior margins are convex. Pectoral length 9.2 (8.8–9.6)% TL and 1.0 times into anterior margin length; pectoral fins much smaller than pelvic fins; pectoral origin is posterior to sixth gill opening. Pectoral fin skeleton radials (SAM 36076) extend about 3.6 times into length of pectoral anterior margin; radials divided into a maximum of six segments ( Figure 3 View FIGURE 3 ). Propterygium small with no radials, mesopterygium with 7 radials, and metapterygium with 4 radials on basal segment and 4 on metapterygial axis; total radial counts were 15 ( Table 3).
Pelvic fins large and broadly rounded, with anterior margin 1.0 times pectoral anterior margin; anterior and posterior margins convex; inner margin sexually dimorphic with males being somewhat longer than females. Inner margin of male paratypes do not form a clasper scroll. Pelvic fin skeleton (SAM 36076) is a flattened band of cartilage, slightly concave dorsally and enlarged at the ends; a long basipterygium with three segments from which 20 radials extend diagonally from its axis ( Table 3). Pelvic radials with four segments each, except for the last two that are unsegmented.
The claspers from one of the male paratypes (SAM 36076) consist of an elongated axial cartilage connected to the basipterygium by an intermediate segment followed by the beta cartilage. In the adult condition the end-style is joined by a terminal cartilage.
Anal fin very large, broadly rounded, its height is 2.0 times dorsal fin height, base length 1.7 in dorsal base. Anterior and posterior fin margins are rounded and convex; an acute angle forms at the tip of the posterior and inner margins. Fin height 2.3 to 2.7 times into base and 2.0 to 4.2 times inner margin. Anal fin skeleton (SAM 36076) composed of an elongated basal cartilage with 30 radial cartilages ( Table 3) extending diagonally from it; each radial has 4 segments.
The dorsal fin is set far back, about 64.2 (62.6–64.5)% TL from snout tip, and low, only 1.9 times anal fin height. Anterior margin is rounded and convex with posterior margin. Base is short, 0.5 times dorsal-caudal space, height 3.1 times in base, and inner margin 0.9 times in height and 3.1 in base. The dorsal fin skeleton (SAM 36076) has a basal cartilage that is elongate, slightly convex distally and flattened proximally. Basal cartilage radials number 15 with each having four segments ( Table 3).
Caudal fin elongated, subtriangular, and without a subterminal lobe; length of dorsal margin 2.6 times in precaudal length.
Total vertebral column count from SAM 36076 was 147. The MP-DP counts were 18 MP and 76 DP for a total precaudal count of 94 ( Table 4). The transition between the MP and DP vertebrae occurred just posterior to the pectoral fins at the 18th vertebra. The caudal count (DC) was 53.
The cranium of one paratype (SAM 36076) was dissected out and measured ( Table 5). Neurocranium ( Figure 4 View FIGURE 4 ) short, with a blunt rostral cartilage, not hypercalcified in adult. Medial rostral cartilage length is about 19 times into nasobasal length. Width across lateral rostral cartilage bases about 4.9 times in length of medial rostral cartilage. Nasal capsules longer than wide, with rounded edges. Width of cranium across nasal capsules 1.7 in nasobasal length. Nasal apertures moderately large and transversely oval, separated by a space 1.8 times there widths. Anterior fontanelle large, rounded anteriorly, sub-triangular posteriorly, slightly longer than wide. Cranial roof 1.3 times orbital length. Orbital notches 1.3 times orbital length. Suborbital shelves 0.7 times orbital length. Otic capsules 0.9 times orbital length.
The number of spiral valve turns was 27 for the holotype and ranged from 26–28 for the paratypes ( Table 6).
The liver is large and extremely oily, consisting of two lobes, one on each side of the body, extending the entire length of the abdominal cavity. The lobes are of equal size and alike in form.
Coloration. In life a dark chocolate brown, but with a thin membranous mucous covering the shark that upon being rinsed off after death revealed a dark gray color beneath the membrane. Color after preservation varies from light brown to gray.
Etymology. The species name africana is in allusion to the Marine and Coastal Management research vessel “ Africana ” in recognition of the excellent research surveys this vessel has conducted over the past several decades. It is also the vessel that collected the paratype SAM 36076.
Distribution. In the southeastern Atlantic, C. africana is confirmed only from off southern Angola southwards to southern Namibia ( Figure 5 View FIGURE 5 ). Although Indian Ocean records are scarce, Chlamydoselachus species have been caught in oceanic waters off the Transkei coast, Eastern Cape Province, South Africa ( Timokhin, 1980) and off the kwaZulu-Natal coast (R. White, fishermen, kwaZulu-Natal, South Africa, pers. comm.). Specimens caught from off the Transkei coast were taken in depths of between 1230 and 1400 m ( Timokhin 1980), while the kwaZulu-Natal specimen was caught in about 300 m depth by a ski boat fisherman (R. White, pers. comm.). One additional Eastern Cape record worth noting was reported by Smith (1951) of an angler who supposedly caught a Chlamydoselachus species from off a pier at Port Alfred.
Unfortunately, the specimen was discarded before its identity could be confirmed. One of us (DAE) spoke with Margaret M. Smith (now deceased), former director J.L.B. Smith Institute of Ichthyology, Grahamstown, South Africa in December 1986 who knew the angler personally that caught this specimen and considered him to be very reliable in fish identification. Given that the continental shelf is very broad in this area it seems unlikely that this deepwater species would occur so far out of its normal range. It may be that this particular specimen was ill, disabled, or at least disoriented when caught. Thus, this record is only tentatively included here. Identification of these latter specimens as to whether they represent the new species, C. africana , or C. anguineus is unknown as these specimens were unavailable for examination.
Size. The largest female on record, an immature specimen, is the holotype that measured 1170 mm TL. Males reach a maximum length of at least 990 mm TL.
Biological notes. Based on the degree of calcification of the terminal cartilage elements it is concluded that male southern frilled sharks mature at a minimum size of 915 mm TL. The epididymis of adult male C. africana were enlarged and coiled and sperm could be expelled by applying pressure along the posterior portion of the genital papilla. Tanaka et al. (1990) concluded that the onset of maturity for male C. anguineus occurred at approximately 1100 mm TL. The smallest adult male recorded by Tanaka et al. (1990) was 1178 mm TL. Claspers of the Chlamydoselachidae are not enclosed within a scroll formed by the pelvic fin inner margin as in the Hexanchidae , but a hexanchid-like clasper sac is present in adult males of both C. africana and C. anguineus .
No adult female C. africana were examined during this study. However, it is assumed that development is viviparous without placenta. Litter size is at least three based on an observation made by Dr. A.I.L. Payne (pers. comm., Cefas, Suffolk, England) of a specimen caught off Namibia, but not retained. Maturity in female C. anguineus has been variously reported from 1256 mm TL ( Tanaka et al., 1990) to approximately 1500 mm TL ( Gudger, 1940). A 1398 mm TL specimen illustrated by Gudger (1940) was immature while a 1550 mm TL specimen was mature. Tanaka et al. (1990) in an extensive study of C. anguineus found the minimum size of maturity in females to be 1256 mm TL. Fecundity has been reported as between 2 and 12 ( Gudger, 1940; Tanaka et al., 1990).
Examination of the holotype and paratypes of C. africana yielded some dietary information on this species. One paratype (SAM 36076), measuring 916 mm TL, contained a partially digested Apristurus sp., measuring approximately 265 mm from the cranial base to the caudal region and weighing 98.8 gm. Another paratype ( RUSI 2424), 1010 mm TL, contained the skeletal remains of a digested cartilaginous fish, possibly a member of the family Scyliorhinidae . A 990 mm TL specimen (USNM 203466) from southern Angola had a squaloid spine embedded in its stomach. The 1170 mm TL holotype (SAM 31028) had a nearly intact Galeus polli measuring 342 mm TL and weighing 97.7 gm in its stomach. On the basis of the limited available data, it would appear that both species of frilled sharks prey primarily on scyliorhinid sharks though they probably feed on other items of an appropriate size. Taniuchi (1987) reported the remains of an Apristurus japonicus in a Japanese specimen. A North Atlantic specimen examined during this study was also found to contain the remains of an Apristurus sp. in its stomach. Unfortunately, very little else is known about the feeding habits of Chlamydoselachus species to confirm their suspected prey preference. However, predation on scyliorhinids and possibly squaloids, as well as other smaller deepwater sharks, would coincide with suspected movement patterns of both predator and its potential prey. Evidence has shown that Chlamydoselachus species are commonly caught in midwater trawls ( Shiobara et al., 1987). Similarly, members of the genera Apristurus and Parmaturus are known to migrate vertically into the water column ( Lee, 1969; Cross, 1988; Ebert et al., 1996).
TABLE 7. Hydrographic data for C. africana (SAM 36076) caught off Namibia at 26º 38' S, 15º 10' E on 19 February 1988. Key: salinity, S o/oo; dissolved oxygen, O 2; nitrogen, N; silicates, Si; phosphates, P; chlorophyll A, Chl A.
The epipelagic ommastrephid squids found in the stomachs of Suruga Bay C. anguineus are fastswimming, highly active and relatively large species ( Kubota et al., 1991). It was suggested that Chlamydoselachus may not be sufficiently swift to catch healthy fast squids but might prey on spent, post reproductive squids and injured individuals. Unfortunately the squid prey of C. anguineus examined by Kubota and colleagues were highly digested and could not be examined for reproductive condition or injuries. Chlamydoselachus species may indeed select spent or injured squids, but the possibility also exists that perfectly healthy epipelagic squid could fall prey to these sharks. The long mouth of the Chlamydoselachus species can accommodate relatively large prey, and their snakelike heads and firm, muscular bodies suggest that they may be able to slowly approach fast epipelagic squids, or might be approached closely by squid investigating or attacking them, and make a sudden snakelike lunge to snag them with their relatively strong, tooth-studded jaws. The mouth of Chlamydoselachus bears a functional resemblance to a squid jig, with many needle-sharp, inward and diagonally-directed curved points on its teeth which is enhanced by the outward rotation of the tooth rows when the jaws are protruded. Even a glancing strike by a shark on a squid or other soft-bodied prey could readily snag it. Squid might also jig themselves by grabbing at the conspicuous whitish tooth rows. Live Chlamydoselachus have been photographed swimming in captivity with mouth agape and with tooth bands highlighted by the dark mouth, but whether this lures squids into striking at the teeth and hooking themselves is speculative at present.
Examination of fresh caught specimens of C. africana from southern Africa and C. anguineus from Taiwan revealed that the jaws and buccal cavity are highly distensible, suggesting that these sharks are capable of ingesting quite large prey items. The inwardly projecting, needle-sharp teeth further indicate that this is a highly specialized feeder. In addition, the abdomen is elongate and the stomach distensible, further indicating that this predator is capable of ingesting fairly large prey. Given these morphological characteristics, e.g. the elongated abdomen, anguiform tooth arrangement, terminal mouth, and highly distensible jaws, these sharks appear to have a rather specialized body arrangement reminiscent of gulper eels ( Saccopharyngidae ) or viperfish ( Chauliodontidae ). Furthermore, Chlamydoselchus species are capable of swallowing relatively large prey items ranging from 1/3 to at least 1/2 their own body length, in much the same way as gulper eels and viper fish are able to engulf large prey items. The Apristurus sp. found in the 916 mm TL Namibian specimen measured 29% of the predator's TL. However, this was only a partial measurement since most of the head and half the tail region had been digested. Forensic examination of the prey item indicated that it had been ingested whole. In life, it would probably have measured closer to 35 or 40 % of this shark's TL. The unique body shape of the Chlamydoselachidae is optimal for grasping and engulfing prey and their ability to forage in both demersal and midwater habitats indicate a highly evolved life style that is not remotely primitive. Thus, the Chlamydoselachidae appear to be highly specialized predators.
Habitat. Hydrographic and substratum data are virtually unknown for Chlamydoselachus records published to date. Hydrographic data on a paratype (SAM 36076) caught at a depth of 425 m off southern Namibia indicate that it was in a low oxygen, high nutrient zone (Table 7), with a soft bottom substratum.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Chondrichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Chlamydoselachus africana
| Ebert, David A. & Compagno, Leonard J. V. 2009 |
Chlamydoselachus anguineus:
| Smith 1951 |