
Tyranniphilopterus polioptilus, Cicchino, Armando C., 2007
|
publication ID |
https://doi.org/ 10.5281/zenodo.178014 |
|
DOI |
https://doi.org/10.5281/zenodo.6251916 |
|
persistent identifier |
https://treatment.plazi.org/id/F320DB6E-FFB2-FFCF-FF71-6B4DFF4DFDAE |
|
treatment provided by |
Plazi |
|
scientific name |
Tyranniphilopterus polioptilus |
| status |
sp. nov. |
Tyranniphilopterus polioptilus , new species
( Figs. 1–16 View FIGURES 1 – 4 View FIGURES 5 – 12 View FIGURES 13 - 17. 13 - 16 )
Type host: Polioptila dumicola dumicola (Vieillot, 1816) . This species has a wide geographical range, from NE Brazil (States of Pará, Goiás, Minas Gerais, São Paulo, Mato Grosso, Paraná and Santa Catarina), Paraguay and Bolivia to Uruguay and Argentina to level of NE Buenos Aires and La Pampa Provinces ( Sick 1997, Narosky & Izurieta 1987, Rohling Ghizoni, Jr 2004.
Male: ( Fig. 1 View FIGURES 1 – 4 ). Pterothorax with 7–8 setae each side. Abdominal chaetotaxy: tergocentral setae: II 6 (plus anterior pair), III–IV 8, V 6–8, VI–VII 8, IX 6; sternal setae each side: II 1–2 (small), III–V 2 (one long, one short), VI 2 (both long); paratergal setae: II–III 0, IV–VIII 2. Genitalia: parameres long, approximately of the same length as the whole mesosomal complex, best seen in Figs. 2–3 View FIGURES 1 – 4 . Body measurements: AS 3 0.124–0.143 (0.134), POW 0.286–0.295 (0.288), OW 0.362–0.391 (0.379), HL.0.381–0.405 (0.397), PL 0.138–0.148 (0.143), PW 0,214–0.229 (0.218), PTL 0.152–0.171 (0.159), PTW 0.309–0.319 (0.315), AL 0.590–0.619 (0.607), AW 0.433–0.476 (0.444), GL 0.100, GW 0.056, TL 1.129.–1.191 (1.170).
Female: ( Fig. 4 View FIGURES 1 – 4 ) Much as for male, except in dimensions, usual sexual dimorphism and details of chaetotaxy in some abdominal segments. Abdominal chaetotaxy: tergocentral setae: II 6 (plus one anterior pair), III– VI 8, VII–VIII 6, IX 4. Paratergal and sternal chaetotaxies as for male, except for additional long seta occasionally on II or IV. Vulva with 3–4 medium setae each side ( Fig. 12 View FIGURES 5 – 12 ). Measurements: AS 3 0.143–0.162 (0.154), POW 0.310–0.324 (0.317), OW 0.402–0.414 (0.409), PL 0.154–0.171 (0.168), PW 0.233–0.243 (0.236), PTL 0.143–0.171 (0.159), PTW 0.323–0.348 (0.336), AL 0.723–0.781 (0.754), AW 0.457–0.495 (0.480), TL 1.324–1.405 (1.368).
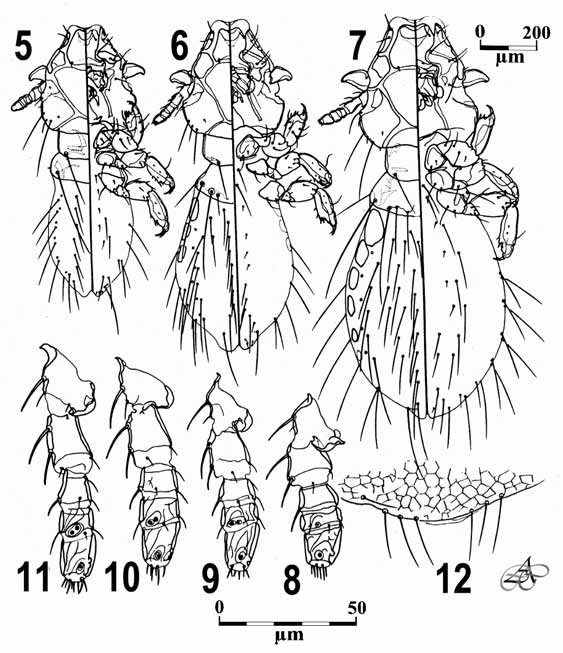
Third nymphal instar: ( Fig. 5 View FIGURES 5 – 12 ). Antennal segment IV with two placoid sensilla arranged in oblique line, similar to adult ( Figs. 10 and 11 View FIGURES 5 – 12 ). Dorsal marginal carina internally notched near middle. Pterothorax with five medium to long posterior setae. Abdomen with sclerotized lightly pigmented paratergal plates present on II– VII. Abdominal dorsal segments II with 4 setae each side, III–V with 3 tergocentral setae, and VI–VII with 2; ventral segments II–VII with 2 setae each side, with innermost small on II–V. Paratergal setae: II–III 0, IV– VIII 2. Body measurements: AS 3 0.133, POW 0.294, OW 0.343, HL 0.381, PL 0.133, PW 0.200, PTL 0.114, PTW 0.281, AL 0.571, AW 0.405, TL 1.143.
Second nymphal instar: ( Fig. 6 View FIGURES 5 – 12 ). Antennal segment IV with two placoid sensilla arranged in a straight line ( Fig. 9 View FIGURES 5 – 12 ). Dorsal marginal carina internally notched near middle. Pterothorax with four medium to long posterior setae. Abdomen with sclerotized lightly pigmented paratergal plates present on II–IV. Abdominal dorsal segments II with 4 setae each side, III with 2 and IV–VII with 1; ventral segments II–VII with 1 seta each side, those on II–V small. Paratergal setae: II–III 0, IV–VII 1, VIII 2. Body measurements: AS 3 0.095, POW 0.238, OW 0.286, HL 0.333, PL 0.124, PW 0.171, PTL 0.105, PTW 0.247, AL 0.409, AW 0.333, TL 0.864.
First nymphal instar: ( Fig. 7 View FIGURES 5 – 12 ). As usual for most Ischnocera , antennal segment IV has only one placoid sensillum ( Fig. 8 View FIGURES 5 – 12 ) (see Cicchino & Abrahamovich, 1988), pterothorax with only one very long posterior seta, and abdomen lacking sclerotized paratergal plates. Dorsal marginal carina completely interrupted near middle. Abdominal dorsal segments II–VII with one long seta, ventral ones similar to N II. Body measurements: AS 3 0.092, POW 0.224, OW 0.273, HL 0.285, PL 0.095, PW 0.171, PTL 0.105, PTW 0.219, AL 0.288, AW 0.262, TL 0.743.
Egg: elongated. Amphora noticeably constricted in proximity of callus ( Figs. 13, 14, 16 View FIGURES 13 - 17. 13 - 16 ), and gradually tapering posteriad ( Fig. 13 View FIGURES 13 - 17. 13 - 16 ), its surface being smooth. Operculum lacking in all 16 eggs examined. Measurements: total length (without operculum), 0.465–0.490 (0.485), maximum width of amphora 0.216–0.224 (0,220). Maximum diameter of amphoral opening 0.114.
Sites of oviposition: one, less commonly two eggs are glued to the basal half of the lower barbs of the feather vanes by means of a moderate amount of spumaline ( Figs. 13–15 View FIGURES 13 - 17. 13 - 16 ). Eggs are laid on feathers of pteryla of the throat, mandible, periauriculars, eyebrow and lateral margins of the crown ( Fig. 17 View FIGURES 13 - 17. 13 - 16 ).
Prevalence: low, 11.76%, two individuals parasitized of 17 examined, both from NE Buenos Aires province.
Remarks: Tyranniphilopterus polioptilus n. sp. is the smallest species of the genus described to date. It is morphologically similar to T. titicacae (Eichler and Freund, 1956) and to several undescribed species of this genus parasitic on tyrannid hosts including: Sayornis phoebe (Shaw, 1790) , Empidonax minimus (Baird,W. M. and Baird,S. F., 1843) , Hemitriccus margaritaceiventer (d´Orbigny and Lafresnaye, 1837), Muscisaxicola macloviana (Garnot, 1829) , Myiarchus tyrannulus (Statius Müller, 1776) , Suiriri suiriri (Vieillot, 1818) , Serpophaga subcristata (Vieillot, 1817) , Leptopogon superciliaris Tschudi, 1844 , Leptopogon amaurocephalus Tschudi, 1846 and Hymenops perspicillatus (Gmelin, 1789) in having relatively narrow preantennal margin, anterior seta 3 (as3) stout, rigid and relatively long, parameres long in the male genitalia, and arrangement of the dorsal and ventral abdominal chaetotaxy. It differs from all of them by proportions of the head and dorsal anterior plate, most of the body measurements, and countings of dorsal, ventral and vulvar setae of the abdomen in both sexes.
Mey (2004) recognized two phenotypes based on the size and number of claws of the third pair of legs. All the specimens available of the new species belong to phenotype II by having all legs essentially of similar size and the third pair double-clawed. Mey also pointed out that the clypeal carina is interrupted laterally in the upper third for all nymphal instars, being interrupted or notched in adult stage, generating his types D1 and D2, respectively. Tyranniphilopterus polioptilus sp. nov., together with species parasitic on members of the Elaeninae ( Suiriri , Serpophaga ) and Fluvicolinae (Hymenops) , showed N I with the carina interrupted, N II deeply notched, N III and adult stage notched, showing a regressive status of this character affecting N II and NIII.
The genus Tyranniphilopterus is known to parasitize suboscine passeriform birds belonging to the Tyrannidae , Pipridae , Cotingidae ( Mey, 2004) and Sapayoidae (Cicchino & Barreto, in prep.) in the New World, as well as two families of oscines, Platysteiridae ( Mey, 2004) in the Old World ( Uganda) and Polioptilidae in the New World ( Argentina). The latter includes the subfamily Polioptilinae , with the genus Polioptila Sclater, 1855 , which has 11 described species, ranging from the south of Nearctic to Uruguay and central Argentina in the Neotropics. Until recently (eg. A.O.U., 1998; Sibley & Monroe, 1990), Polioptila has been classified within the Sylviidae or, alternatively, in a single family of its own, Polioptilidae , presumably linked to sylvioid stock of the Old World. Molecular studies using DNA-DNA hybridization techniques concluded that Polioptila fit well within the Certhiidae , being the sister group of the Wrens ( Troglodytidae ) ( Barker, 2004). It is an interesting that Certhiidae in the New World and Troglodytidae (in both Old and New World) are parasitized by species of Philopterus Nitzsch, 1818 ( Mey, 2004; Cicchino & Castro, 1998), whilst Polioptila is parasitized by Tyranniphilopterus . True Certhiidae are parasisized by Philopterus species ( Mey, 2004) and this supports the opinion that the Gnatcatchers ( Polioptilidae ) are not part of the Treecreepers ( Certhiidae ). This agrees with Dickinson (2003), who stated that they belong to their own family, Polioptilidae , and are not closely related to the Certhiidae . Tyranniphilopterus also parasitizes Sapayoa aenigma Hartert, 1903 . This New World species is included in Sapayoidae , a suboscine family cladistically linked to Philepittidae , Eurylaimidae and Calyptomenidae ( Irestedt et al., 2006) , all from the oriental half of the Old World and not reported to be parasitized by Philopterus species.
Modern theories regarding the Gondwanian origin of the perching birds leads me to conclude that Tyranniphilopterus originated in austral Gondwana, parasitizing primarily the tyrannoid stock, and secondarily on several unrelated, but partially sympatric, suboscine and oscine taxa. With the break-up of austral Gondwana, this tyrannoid stock became isolated in the New World. By the end of the Cretaceous the Polioptilidae and Sapayoidae reached the New World by some of the ways suggested by Fjeldså et al. (2003). Others, as Platysteridae, remained in occidental Gondwana. Alternatively, Polioptilidae may have acquired Tyranniphilopterus a posteriori in the New World from tyrannids, cotingas or mankins.
Etymology: the word “ polioptilus ” refers to the generic name of its host.
Specimens examined: all from Buenos Aires province, Argentina: male holotype, 1 male and 4 females paratypes, 3 nymphs III, 1 nymphs II, 1 nymph I and numerous eggs from Punta Piedras, Magalena District, 23-IX-1984, A.C. Cicchino coll.; 2 males paratypes, Laguna Bellaca, Almirante Brown District, I-1991, A.C. Cicchino coll.
Passerine hosts known to be parasitized by Tyranniphilopterus species, including the remaining louse species known from them. (T) indicates type host, taken at species level. (Avian taxonomy as per Dickinson, 2003)


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |