
Betaburmesebuthus Lourenço, 2015
|
publication ID |
https://doi.org/ 10.11646/palaeoentomology.6.1.10 |
|
publication LSID |
lsid:zoobank.org:pub:B822ED53-D559-4DD8-8803-3F2850A00445 |
|
DOI |
https://doi.org/10.5281/zenodo.7757589 |
|
persistent identifier |
https://treatment.plazi.org/id/F308E17F-4351-FFEB-FCCD-48A1FBE41C3A |
|
treatment provided by |
Plazi |
|
scientific name |
Betaburmesebuthus Lourenço, 2015 |
| status |
|
Genus Betaburmesebuthus Lourenço, 2015
Type species. Betaburmesebuthus kobberti Lourenço, 2015 (in Lourenço & Beigel, 2015).
Included species. Betaburmesebuthus kobberti ( Lourenço & Beigel, 2015) , B. muelleri ( Lourenço, 2015a) , B. bidentatus ( Lourenço, 2015a) , B. bellus (Lourenço, 2016) , B. fleissneri ( Lourenço & Velten, 2016a) , B. larafleissnerae ( Lourenço & Velten, 2016b) , B. joergi ( Lourenço & Rossi, 2017) , B. spinipedis ( Xuan, Cai & Huang, 2022) , B. villosus sp. nov., B. fuscus sp. nov., B. pohli ( Lourenço & Velten, 2017) comb. nov. and B. knodelorum ( Lourenço, 2021) comb. nov.
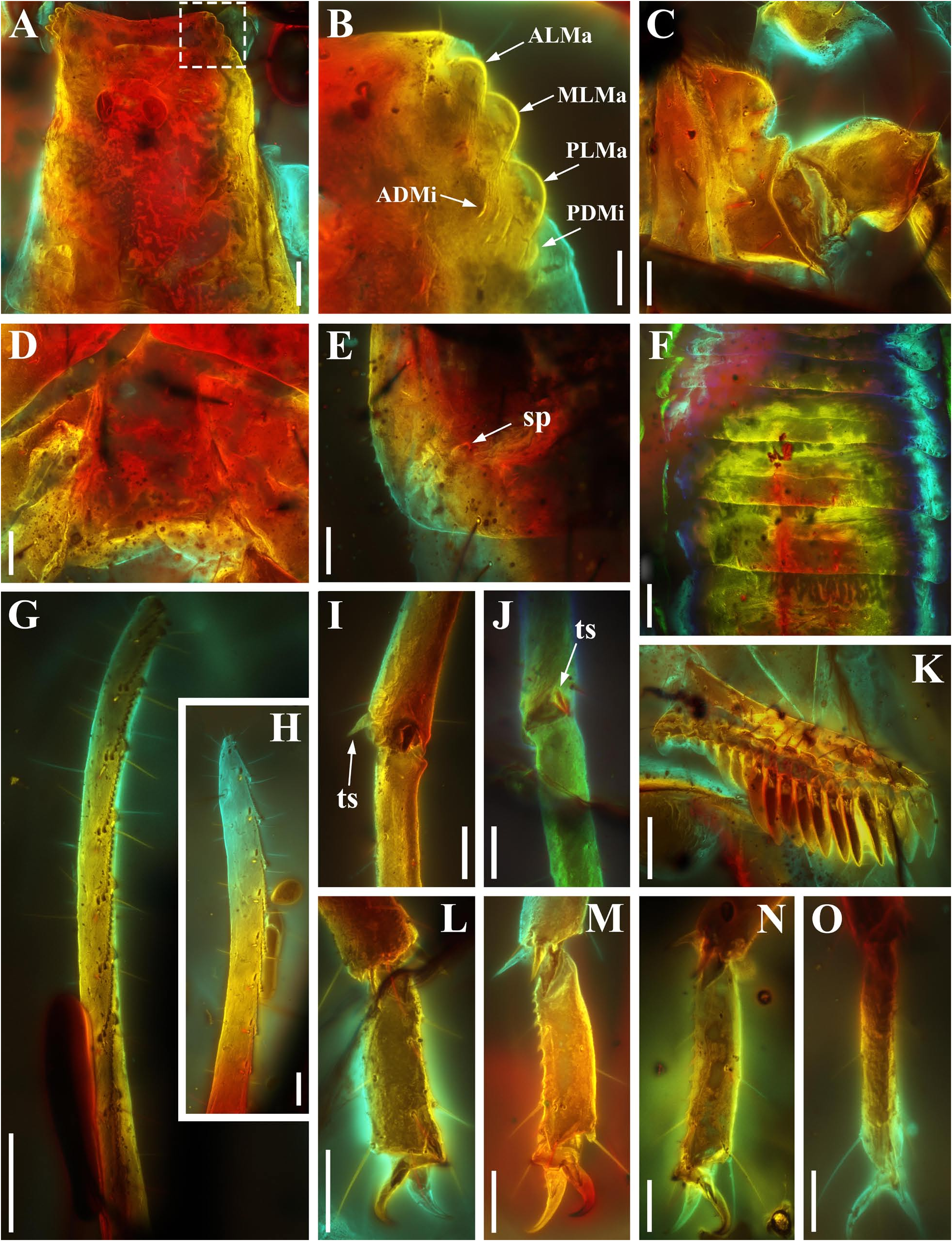
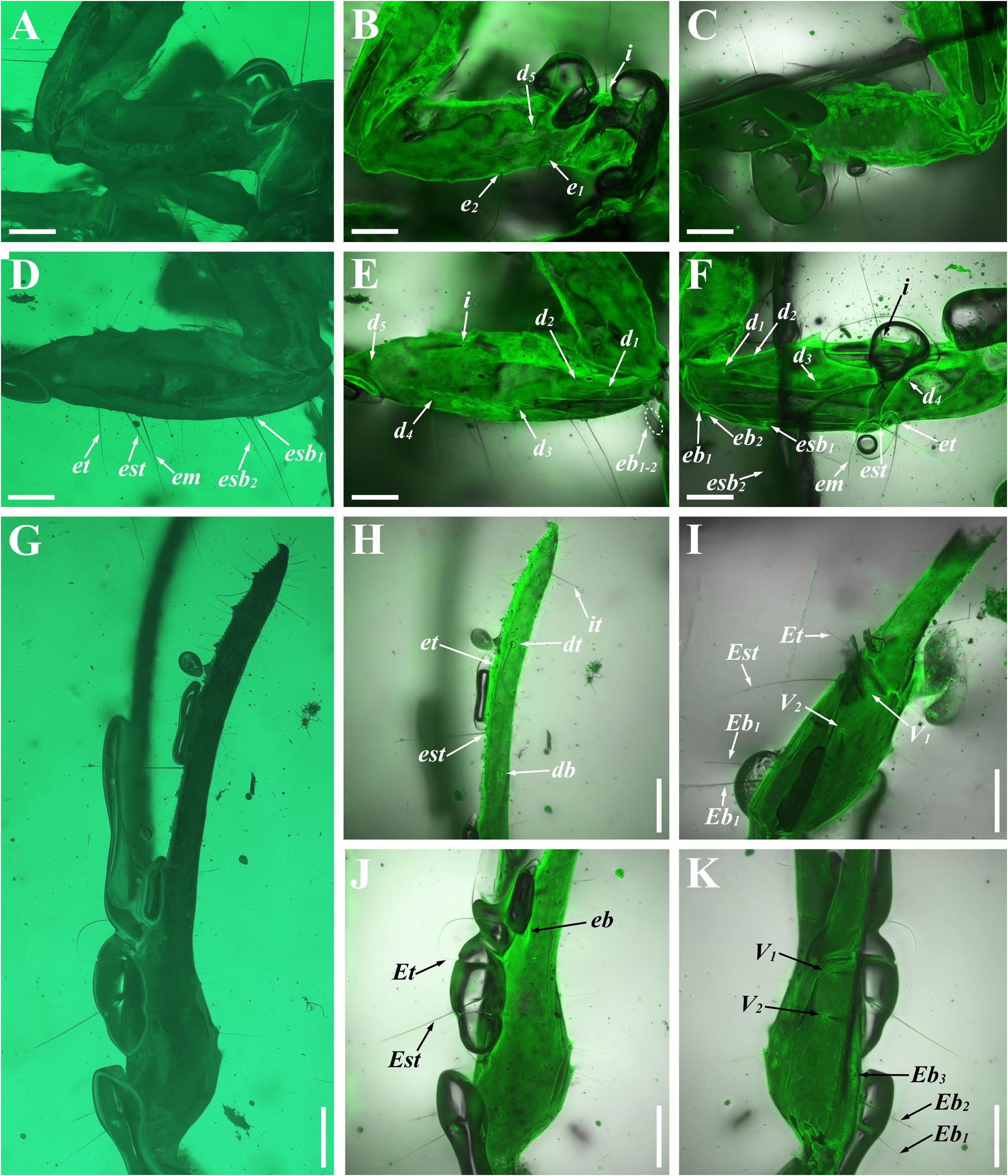
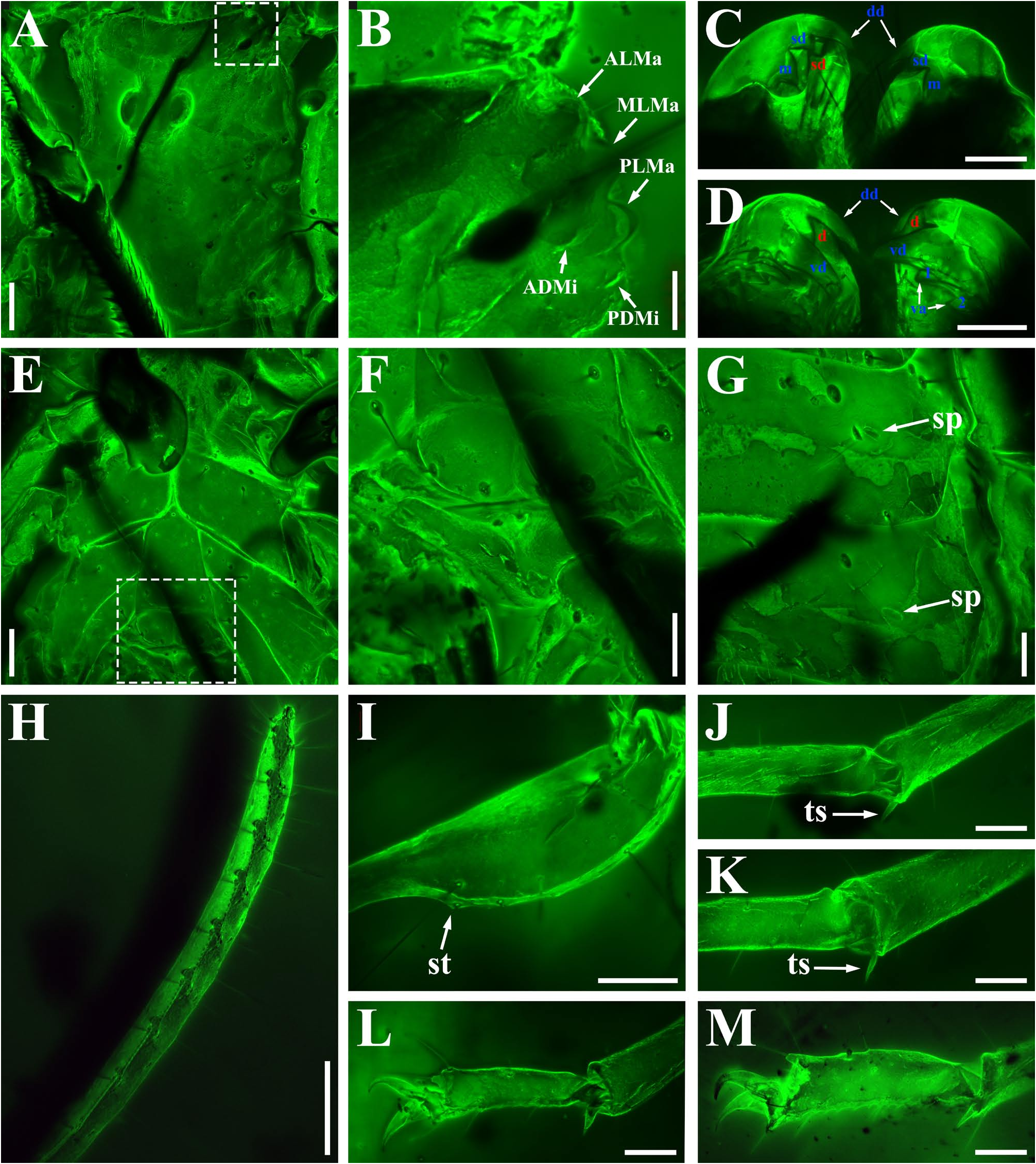
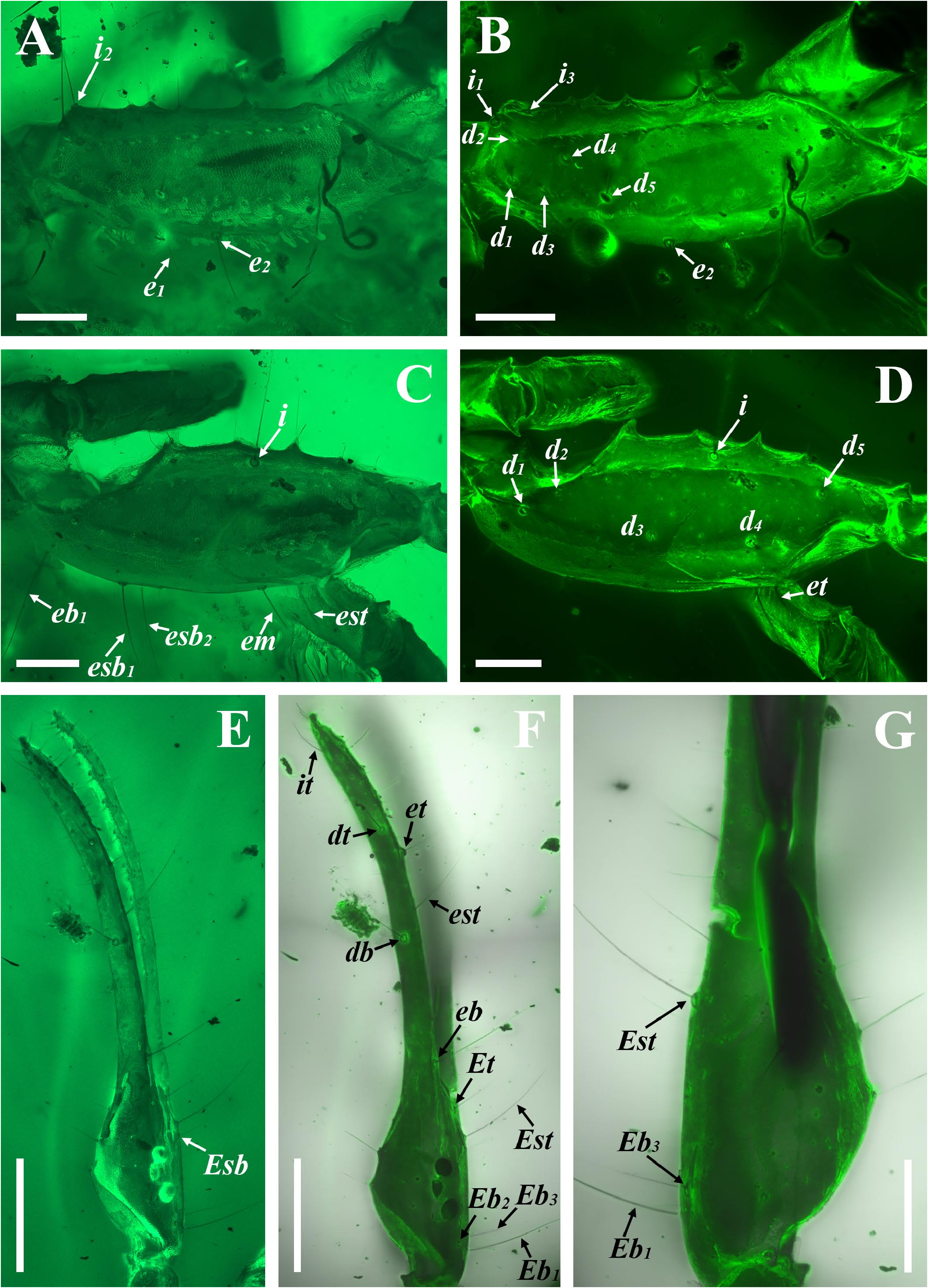
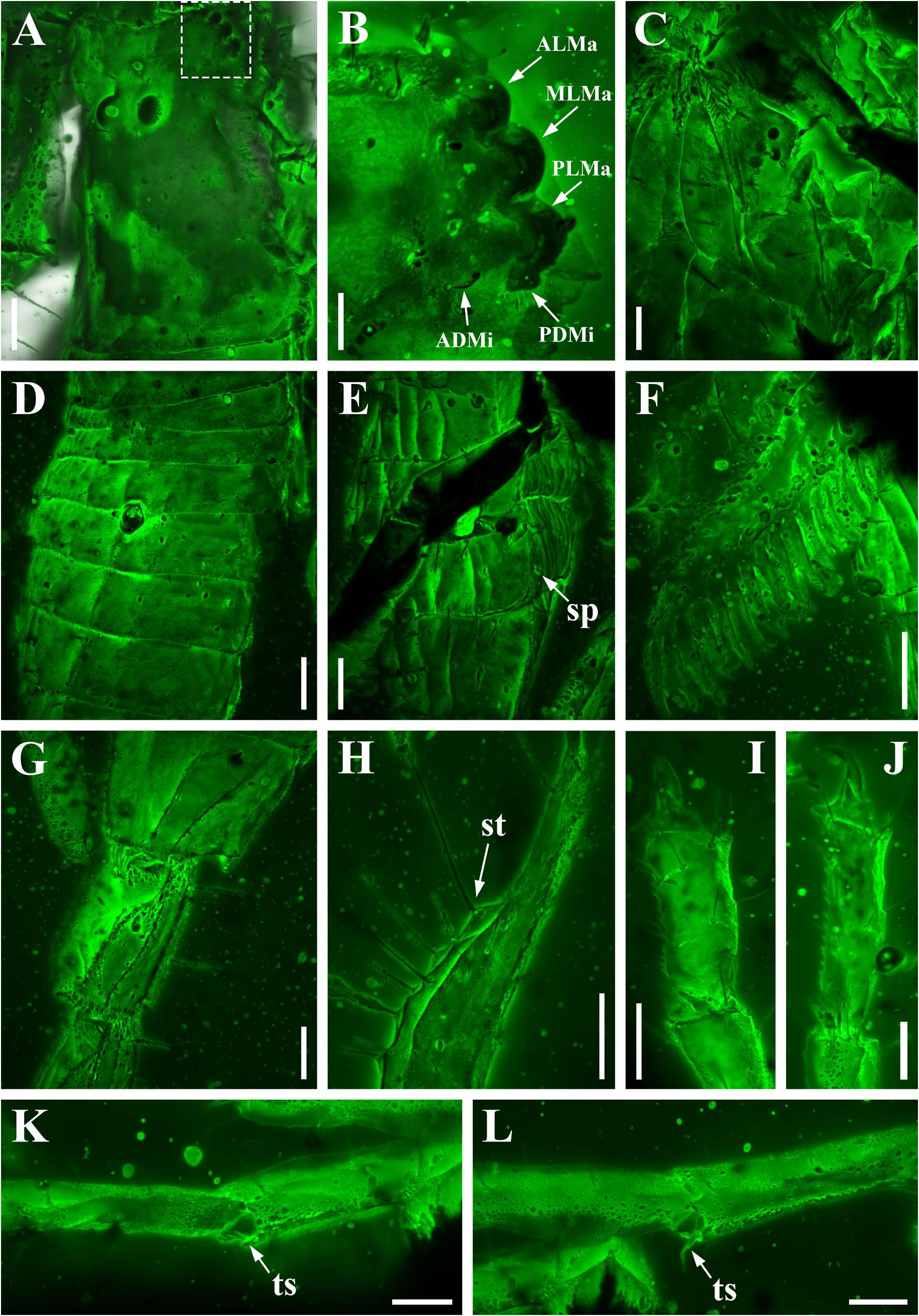
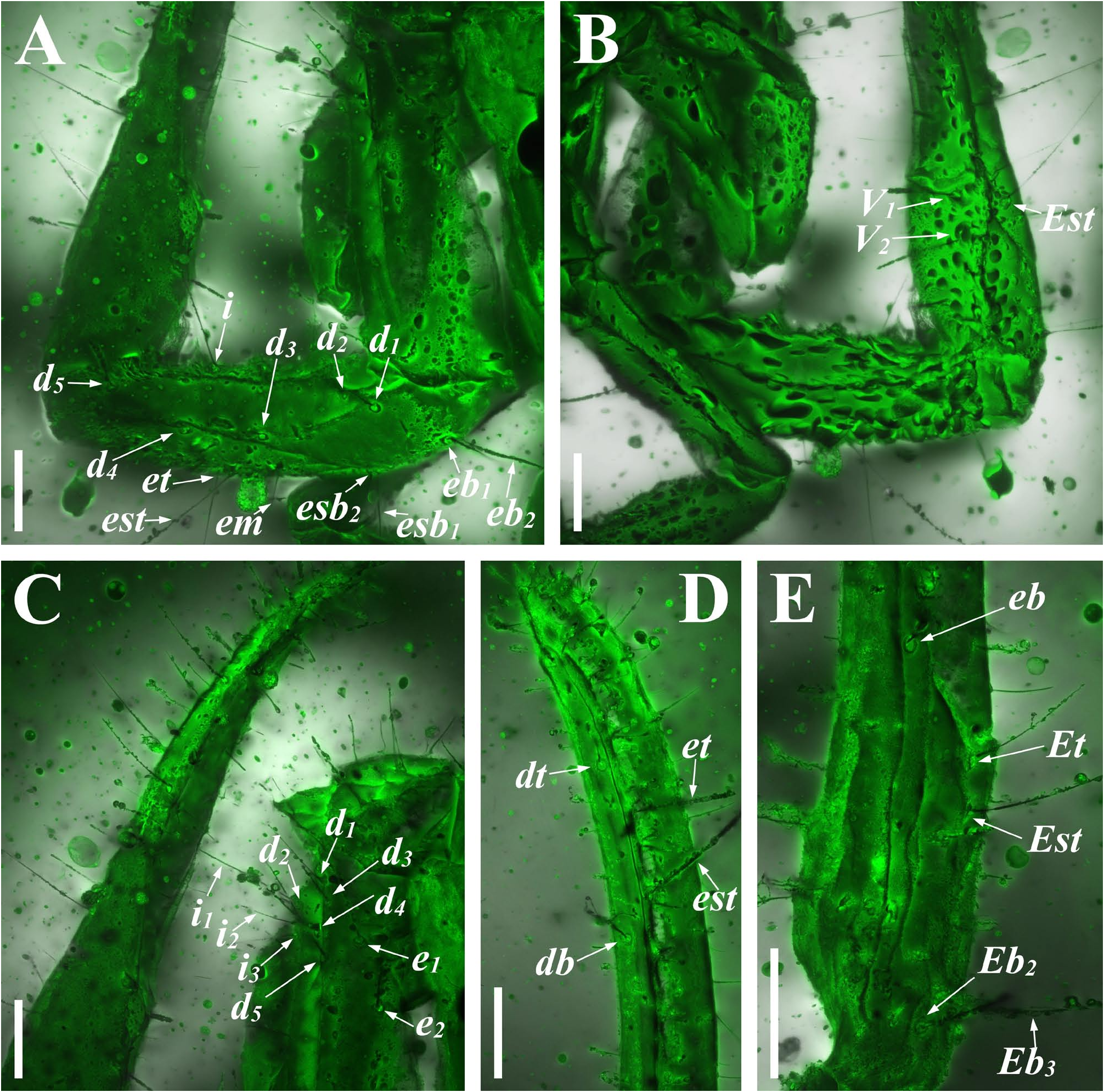
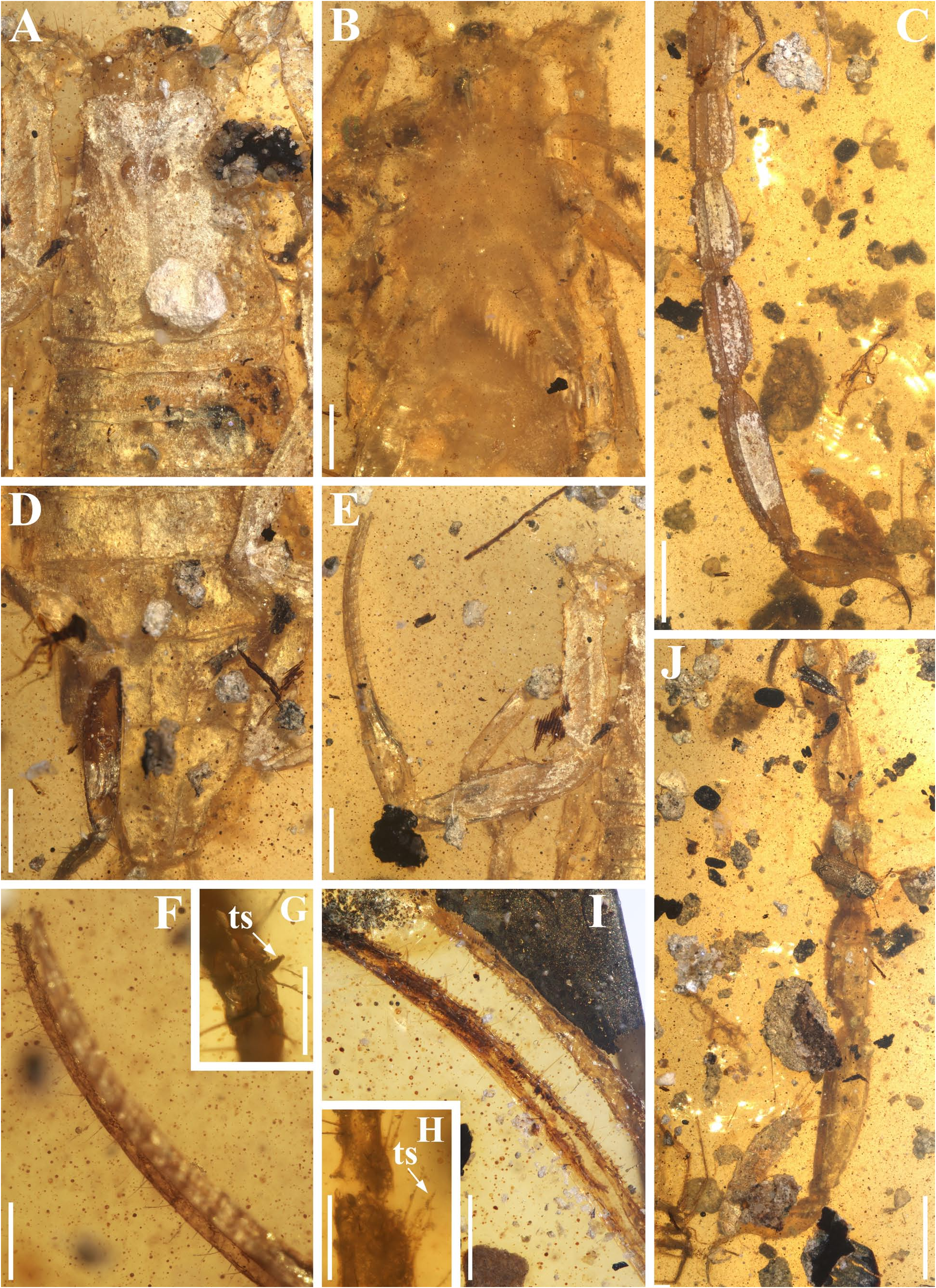
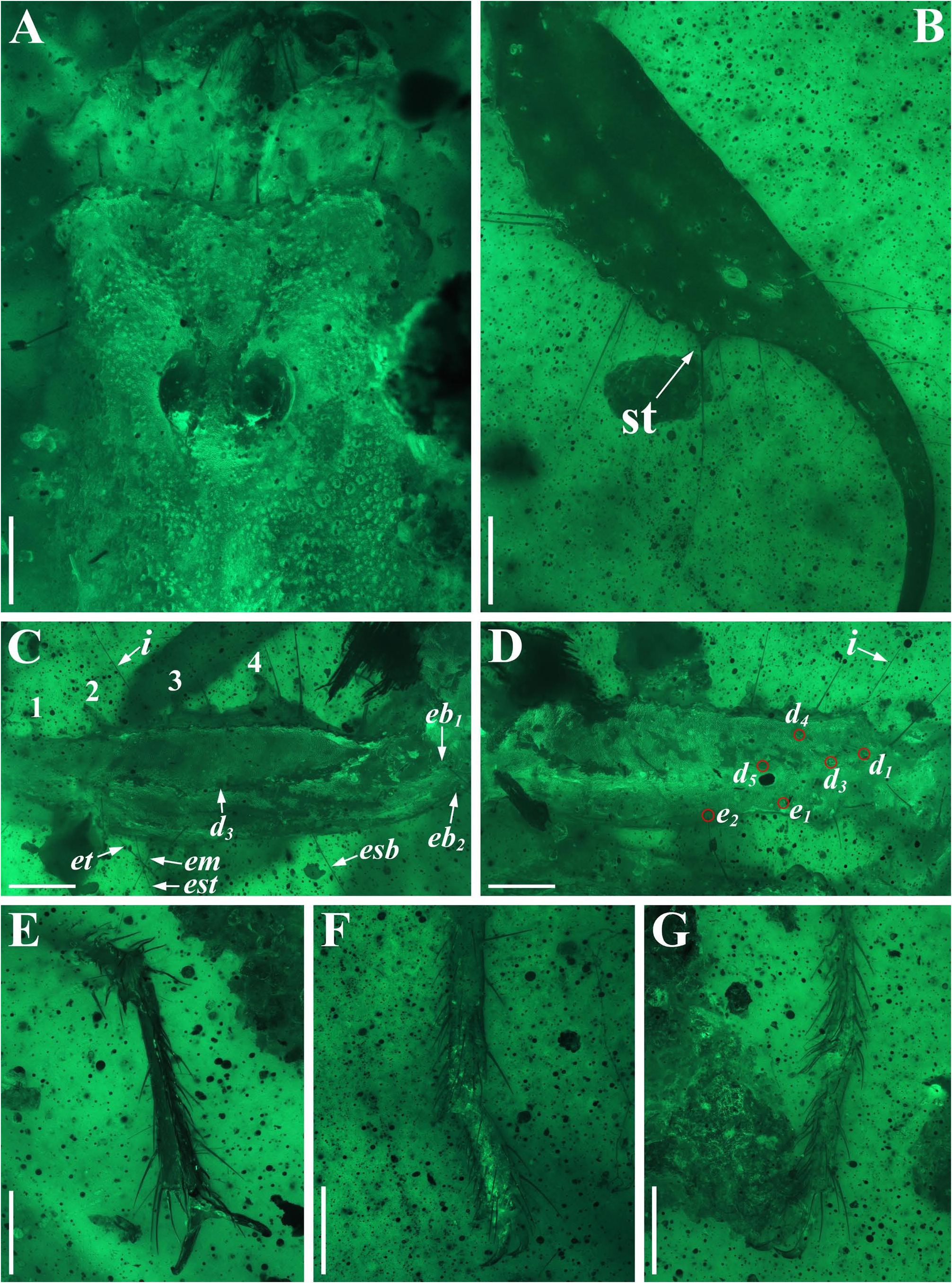
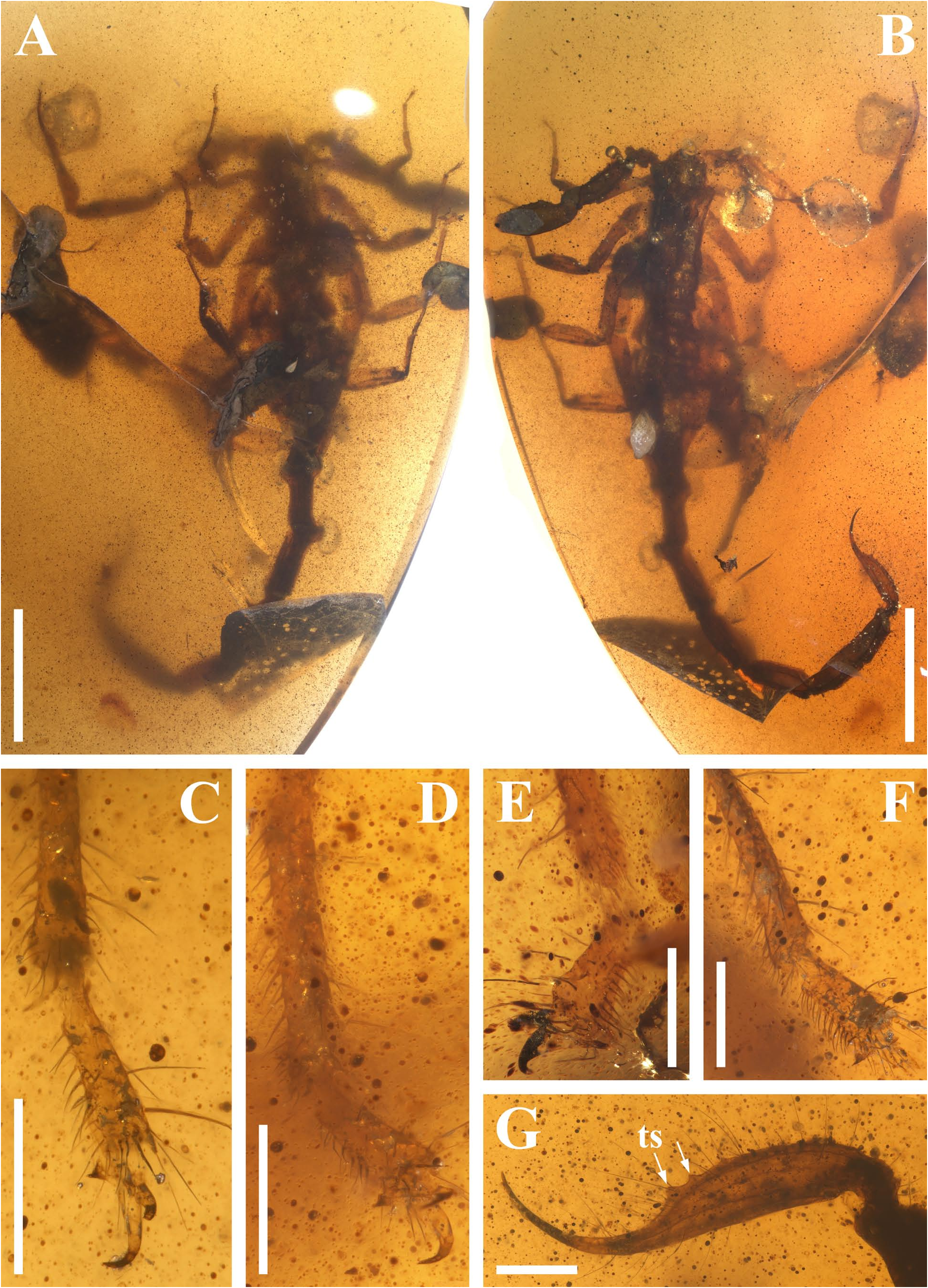
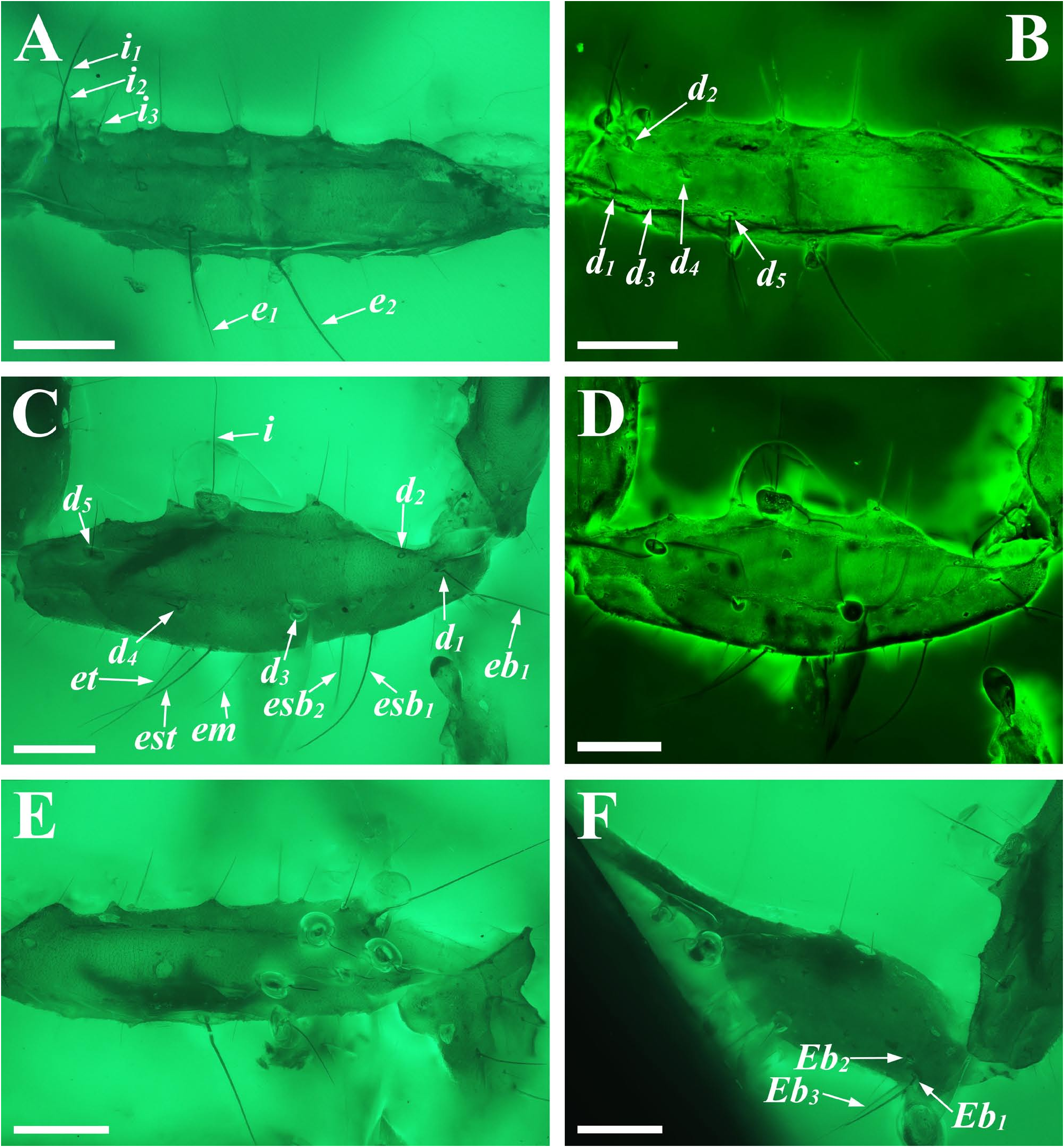
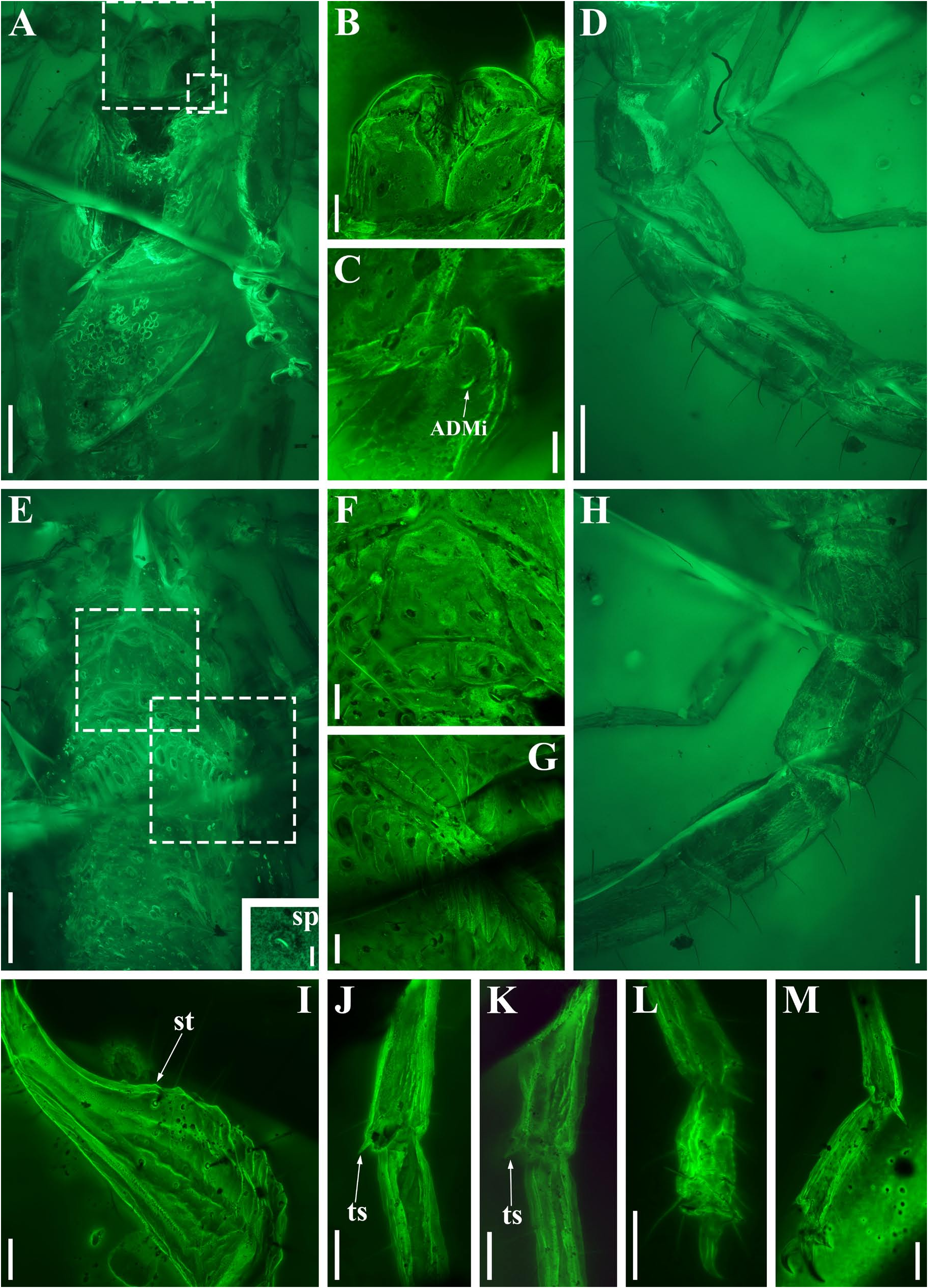
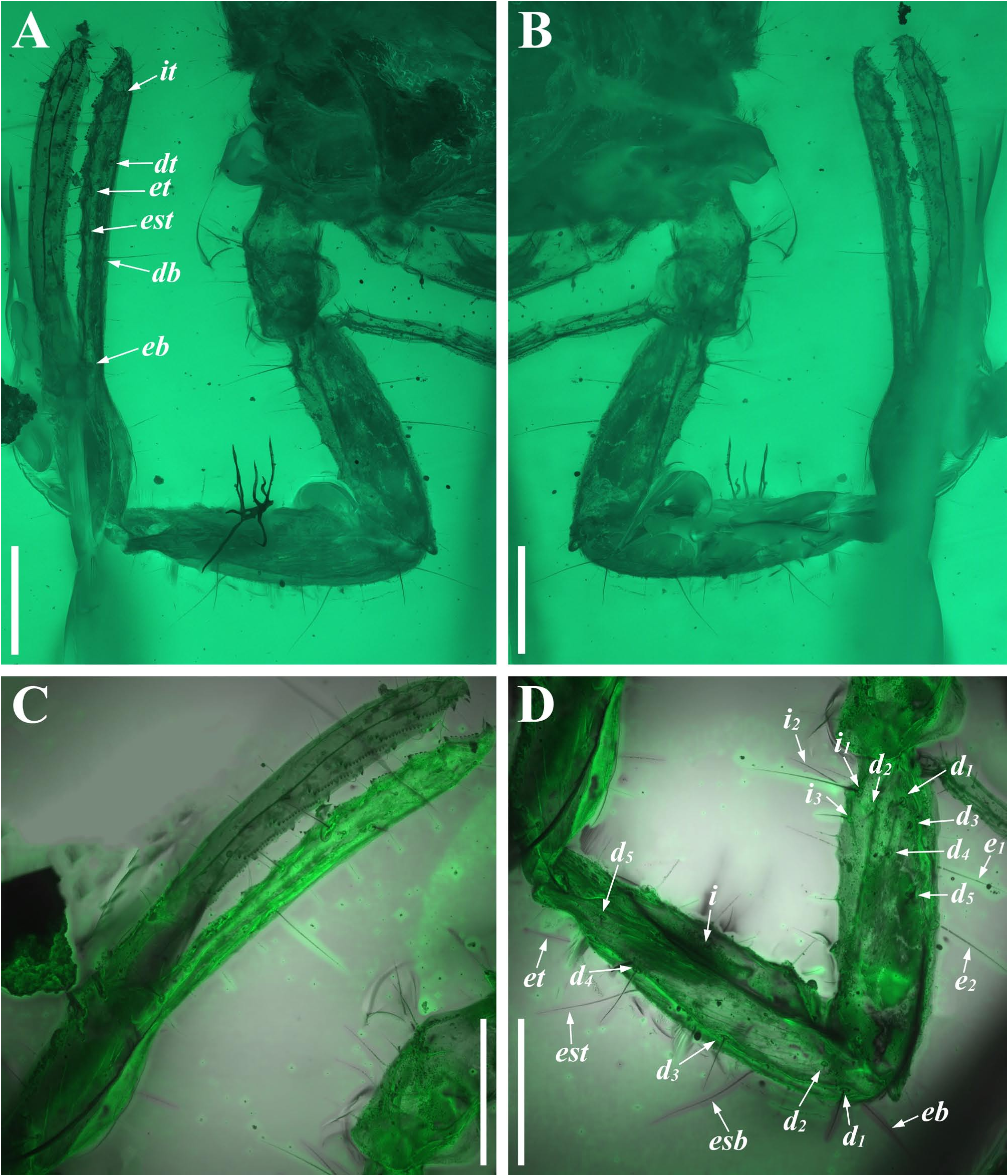
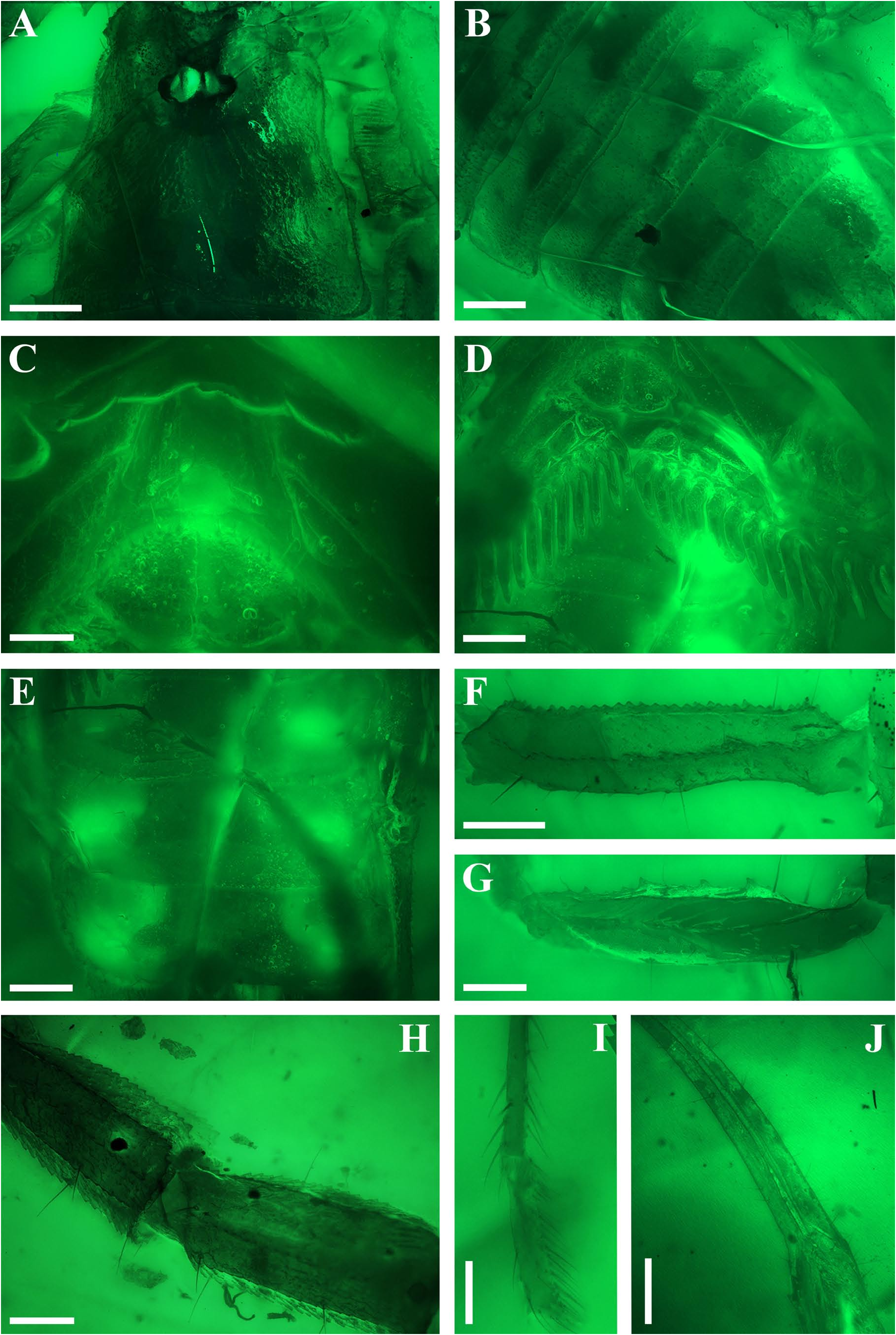
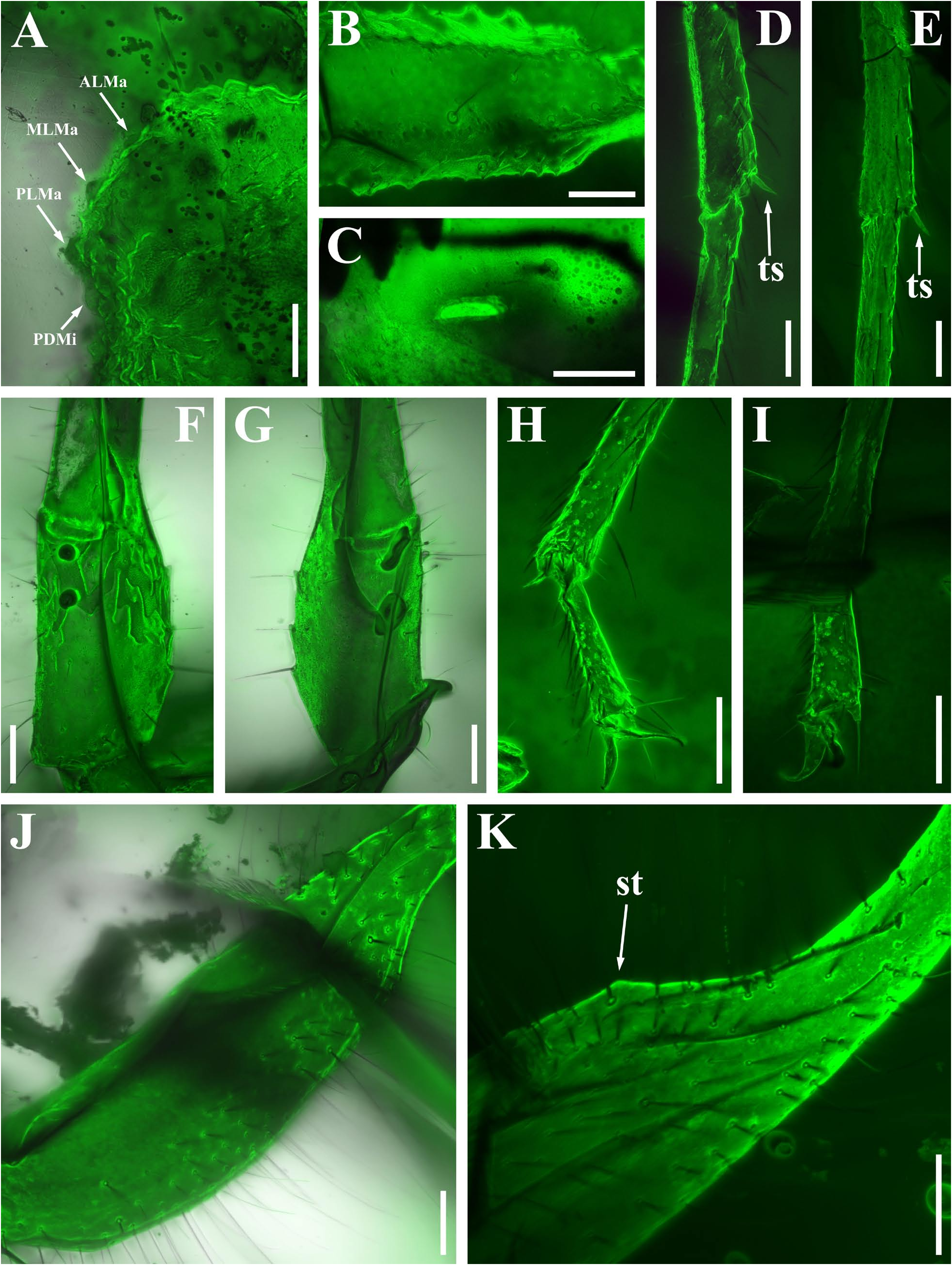
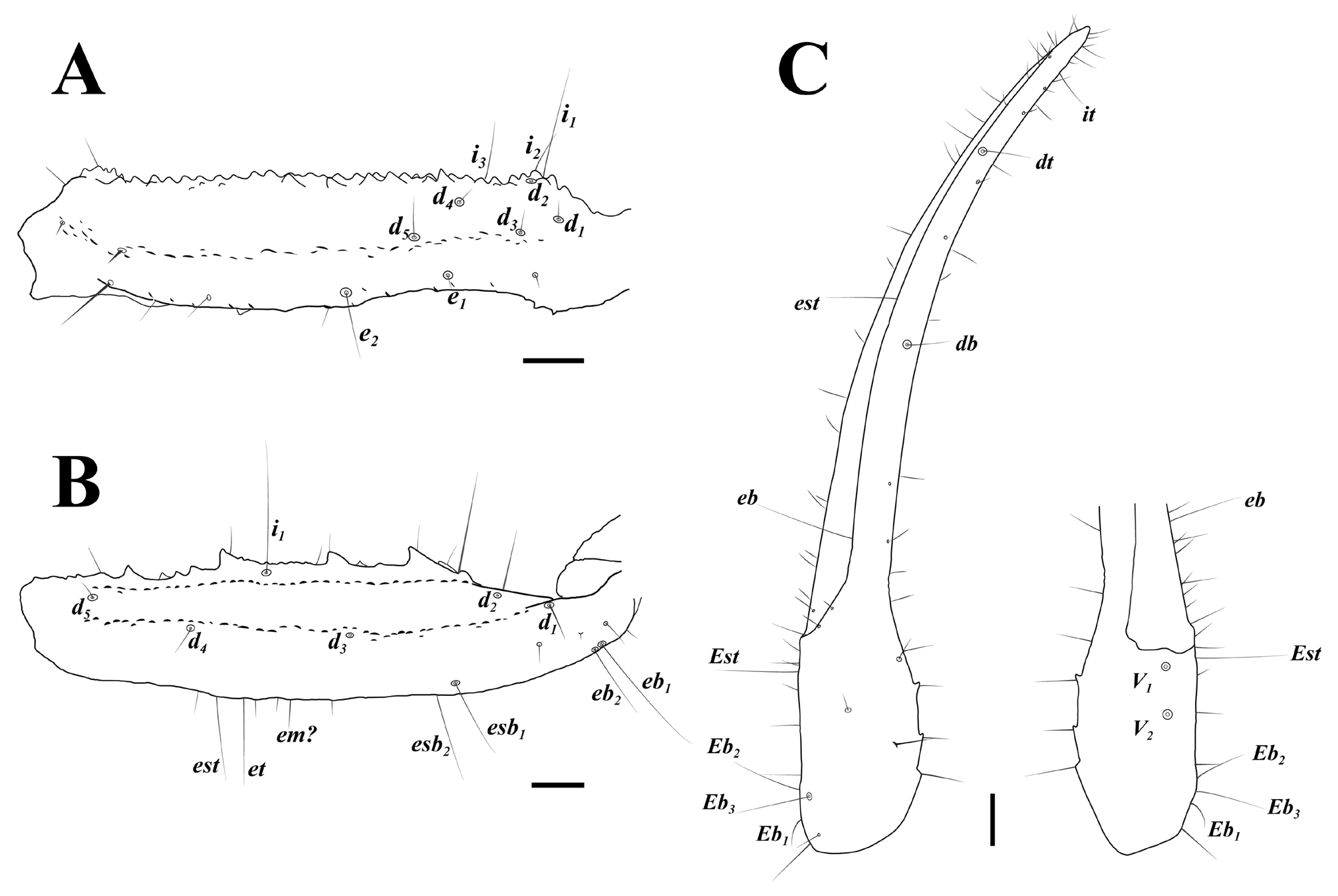
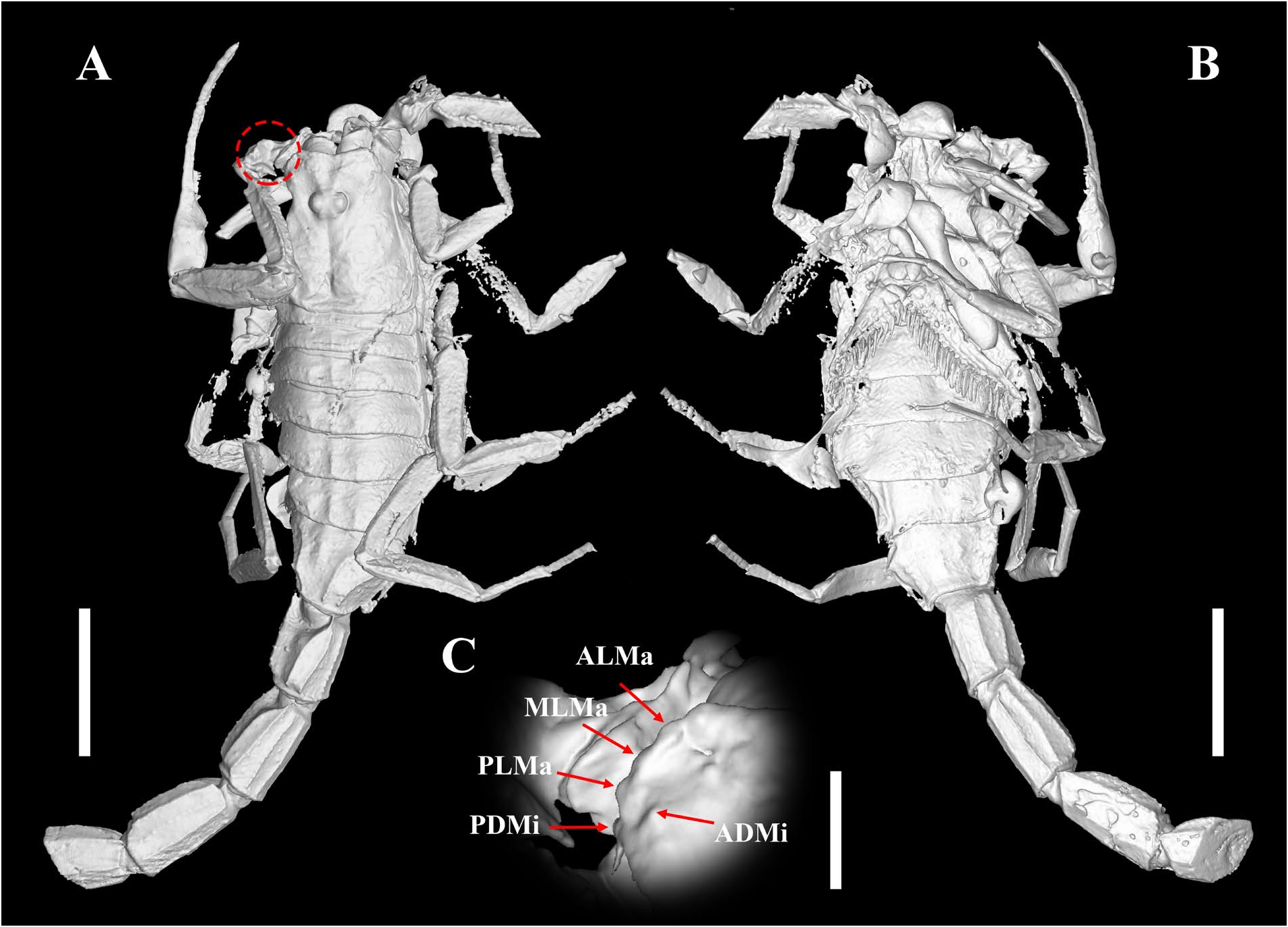
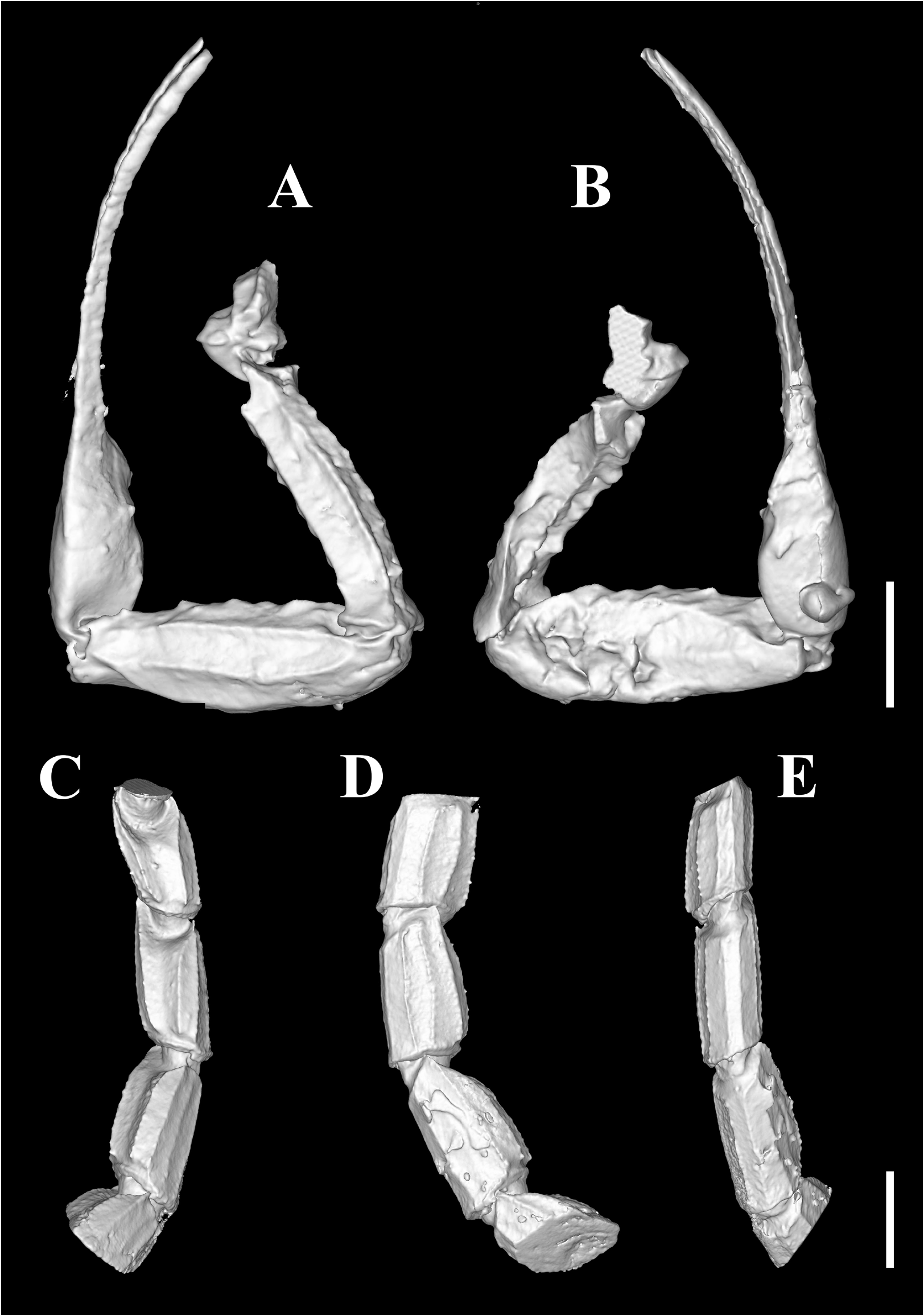
Diagnosis (emended). Five pairs of lateral ocelli present, including three major lateral ocelli (ALMa, MLMa, PLMa) and two minor lateral ocelli (PDMi, ADMi) ( Figs 2B View FIGURE 2 , 6B View FIGURE 6 , 10B View FIGURE 10 , 17B View FIGURE 17 , 20C View FIGURE 20 , 25A View FIGURE 25 , 28C View FIGURE 28 ).Anterior margin of carapace with a wide weakly to strongly marked median concavity ( Figs 2A View FIGURE 2 , 6A View FIGURE 6 , 10A View FIGURE 10 , 14A View FIGURE 14 , 17A View FIGURE 17 , 20A View FIGURE 20 , 24A View FIGURE 24 , 29A View FIGURE 29 ); central median, posterior median and posterior marginal furrows of carapace well-marked in adults ( Figs 13A View FIGURE 13 , 28A View FIGURE 28 , 29A View FIGURE 29 ). Median ocular tubercle well-marked and located on the first anterior third to the first anterior of carapace ( Figs 2A View FIGURE 2 , 6A View FIGURE 6 , 10A View FIGURE 10 , 13A View FIGURE 13 , 17A View FIGURE 17 , 20A View FIGURE 20 , 24A View FIGURE 24 , 29A View FIGURE 29 ). Sternum elongated pentagonal ( Figs 2D View FIGURE 2 , 6E View FIGURE 6 , 20F View FIGURE 20 , 24C View FIGURE 24 ). Pectines with 13–20 teeth and fulcra present ( Figs 2K View FIGURE 2 , 10F View FIGURE 10 , 24D View FIGURE 24 , 29B View FIGURE 29 ). Metasomal ventral carinae parallel ( Figs 5D View FIGURE 5 , 13C, J View FIGURE 13 , 30 E View FIGURE 30 ). Telson with a very long aculeus and a tiny subaculear tubercle ( Figs 6I View FIGURE 6 , 10H View FIGURE 10 , 14B View FIGURE 14 , 17G View FIGURE 17 , 20I View FIGURE 20 , 25J View FIGURE 25 ). Trichobothriotaxic pattern Type A with beta - configuration ( Figs 3 View FIGURE 3 , 7 View FIGURE 7 , 11 View FIGURE 11 , 14C, D View FIGURE 14 , 18 View FIGURE 18 , 21 View FIGURE 21 , 26 View FIGURE 26 ). Pedipalp patellar dorsomedian carina (DMc) well-developed ( Figs 3E View FIGURE 3 , 7D View FIGURE 7 , 11A View FIGURE 11 , 15A View FIGURE 15 , 18D View FIGURE 18 , 21D View FIGURE 21 , 23E View FIGURE 23 , 29D View FIGURE 29 ). Pedipalp finger dentition composed of about ten discrete linear rows of granules; each slightly oblique with proximal ends directed externally, rows slightly imbricated; a smaller spinous external accessory granule and a bigger spinous internal accessory granule present along the base of each denticle row; tip hooked ( Figs 2G, H View FIGURE 2 , 6H View FIGURE 6 , 13F, I View FIGURE 13 , 21A–C View FIGURE 21 ). Tibial spurs present on legs III and IV ( Figs 2I, J View FIGURE 2 , 6J, K View FIGURE 6 , 10K, L View FIGURE 10 , 13G, H View FIGURE 13 , 20J, K View FIGURE 20 , 29F, H View FIGURE 29 ). Telotarsus with few ventrosubmedian setae and a ventromedian spinules row ( Figs 2M, N View FIGURE 2 , 6L, M View FIGURE 6 , 10I, J View FIGURE 10 , 17C View FIGURE 17 , 20L, M View FIGURE 20 ) or numerous ventrosubmedian setae ( Figs 14E–G View FIGURE 14 , 15C–F View FIGURE 15 , 23F View FIGURE 23 , 24I View FIGURE 24 , 25H, I View FIGURE 25 , 29F–I View FIGURE 29 ) .
Remarks. Previous descriptions of Betaburmesebuthus indicated that most members of the genus, except for B. bidentatus , bear three lateral ocelli ( Lourenço & Beigel, 2015; Lourenço, 2015a, 2016; Lourenço & Velten, 2016a, b; Lourenço & Rossi, 2017). Nonetheless, we confirm the presence of five pairs of lateral ocelli in B. spinipedis ( Xuan et al., 2022) . We do not regard this as a new feature since the fourth and fifth pairs of minor lateral ocelli are greatly reduced in size and are likely to be ignored by previous authors. Our examination of studied specimens reveals that B. bellus , B. kobberti , B. fleissneri , B. larafleissnerae , and B. muelleri share this character.
Lourenço & Velten (2017) reported another fossil from Burmese amber, which is assigned to a new genus and species, i. e., Spinoburmesebuthus pohli . Recently, Lourenço (2021) recorded the second species of this genus, S. knodelorum . The most important character of Spinoburmesebuthus claimed by Lourenço (2021) is the presence of subaculear tubercle. However, in another paper Lourenço presented a line-drawing of the adult male metasoma of Palaeoburmesebuthus knodeli ( Lourenço, 2018: fig. 4) which clearly shows a subaculear tubercle. In addition, a small subaculear tubercle also found in the juvenile individual of B. spinipedis ( Xuan, Cai & Huang, 2022) . Thus, this feature become a contradictory identification key, and we examined the additional materials of Betaburmesebuthus in this paper to contribute to the solution of this taxonomic confusion. We conclude that the subaculear tubercle is present in Betaburmesebuthus , which is difficult to discover in immature individuals but distinct in adult individuals. The two species assigned to alleged Spinoburmesebuthus show many similarities with Betaburmesebuthus and are the large specimens, as our materials, B. fuscus sp. nov. (NIGP200653) and B. sp. (NIGP201157), which all exhibit the obvious subaculear tubercle. As a result, we here merge the genus Spinoburmesebuthus into Betaburmesebuthus syn. nov. and transfer the two species of Spinoburmesebuthus to Betaburmesebuthus , i. e., B. pohli comb. nov. and B. knodelorum comb. nov.
The description of denticles of pedipalp fingers for Betaburmesebuthus remains ambiguous, which could be attributed to the fact that the original authors observed this structure only from the lateral view ( Lourenço, 2021: fig. 16). We provide clear fluorescence and confocal photos from the frontal view and suggest that the absence of any conspicuous accessory granules are artifacts. Remarkably, another Burmese amber fossil, Archaeoananteroides maderai ( Lourenço & Velten, 2016c) shows the same state in denticle rows with our studied specimens and this species is likely to be a member of Betaburmesebuthus .
Santiago-Blay et al. (2022) indicated that the tibial spurs III and IV are variable in Betaburmesebuthus (present in three species, absent or reduced in other three species). A detailed examination of our material, however, revealed the presence of tibial spurs III and IV is fixed in members of Betaburmesebuthus . We suggest that such an absent or reduced condition were probably due to the poor preservation or are artifacts of observation. Furthermore, the tibial spurs of immature individuals are usually undeveloped and not easily to identify.
Another interesting character is telotarsus armature. After the examination of all material, we observed two types: few ventrosubmedian setae and a ventromedian spinules row versus numerous ventrosubmedian setae. The feature is rarely mentioned in published papers and should be regarded as a diagnostic character, despite that the telotarsus armature was considered an important taxonomic character in extant scorpion at higher-level taxa ( Soleglad & Fet, 2003). Lacking more distinct diagnostic characters, we refrain from establishing a new genus to divide the two forms.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |