
Cephalodella jersabeki, De Smet, Willem H. & Verolet, Michel, 2016
|
publication ID |
https://doi.org/10.11646/zootaxa.4107.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:EE8363BC-7BE3-4F68-9058-6C3F891BCBBA |
|
DOI |
https://doi.org/10.5281/zenodo.6091369 |
|
persistent identifier |
https://treatment.plazi.org/id/F209BB71-A22B-F30C-FF08-FA50FE966A67 |
|
treatment provided by |
Plazi |
|
scientific name |
Cephalodella jersabeki |
| status |
sp. nov. |
Cephalodella jersabeki n. sp.
( Figs 1 View FIGURE 1 , 2 View FIGURE 2 )
Diagnosis. Female about 160−170 µm long. Body stout, arched dorsally, almost flat ventrally, weakly compressed laterally. Tail rather small, rounded. Toes short, less than 1/5 total length, more or less conical, in dorsal view broadest and arched near base, tapering to rather long, blunt tips; median and ventral margin almost straight; tips weakly recurved dorsally. Two colourless eyespots (?) at end of brain. Trophi type B, moderately asymmetric. Rami composed of three well developed and sclerified chambers; fairly acute and long alulae of almost equal length. Flabella rami small, fan and shaft reduced, composing sclerite elements fused. Pectines rami with left 7 large appressed distal teeth and ~ 4−5 very short appressed proximal knobs, and right 9 large appressed distal teeth and ~ 15−20 short appressed proximal knobs. Manubria straight, characteristic asymmetrical distal crutches; crutch of left manubrium sickle-shaped, outer margin of sickle with short, acute lateral projection; crutch of right manubrium smaller, more or less crooked, outer margin without acute projection; manubrial chambers lamellar, dorsal lamella pointed distally.
Type locality. Canal Font Rome, Aubenas, France; 20 May 2009; epibiotic on G. pulex (L.).
Holotype. A female in a permanent, glycerine glass slide mount deposited in the Royal Belgian Institute of Natural Sciences ( RBINS), Brussels, Belgium, IG 33143, RIR 258.
Paratypes. One female from type locality in RBINS, IG 33143, RIR 259; 3 mounted paratypes and 4 SEM trophi preparations in Department of Biology, University of Antwerp.
Etymology. The species is dedicated to Dr Christian Jersabek in recognition of his rotifer studies and impressive Rotifer World Catalog (http://www.rotifera.hausdernatur.at).
Description of female. Body ( Figs 1 View FIGURE 1 A, B, E) stout; in lateral view dorsal margin gibbous, ventral margin almost straight; in dorsal view weakly compressed laterally, widest near mid-length. Head moderately large, slightly deflexed, offset by distinct neckfold. Integument soft, no trace of lorica plates. Tail rather small, rounded, may reach up to midlength of foot. Foot stout, irregular conical, very slightly deflexed. Toes ( Figs 1 View FIGURE 1 C, D) short, less than 1/5 total length, more or less conical, in dorsal view broadest and arched near base, tapering to rather long, blunt tips; medial and ventral margin almost straight; tips weakly recurved dorsally; proximal part of toes with reservoir, separated from distal duct by minute light refracting structures. Corona fairly oblique. Eyespots (?) two colourless globules at end of brain. Dorsal antenna in posterior third of head. Brain large, saccate, extending till neckfold. Proventriculus present. Gastric glands large, short-stalked, lobed. Stomach large, indistinctly separated from short intestine. Pedal glands large, extending into trunk, with small accessory glands (?). Vitellarium with 8 nuclei.
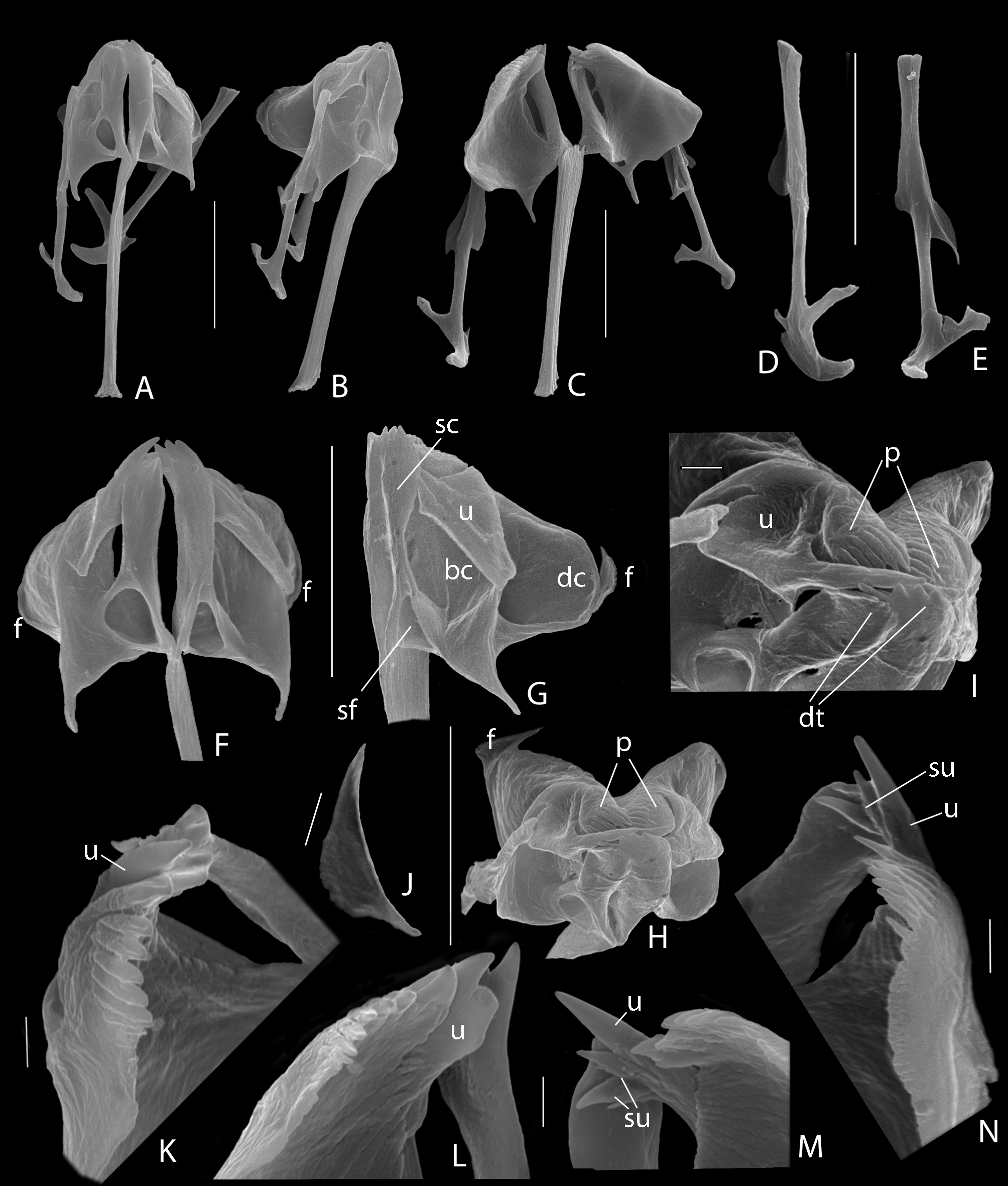
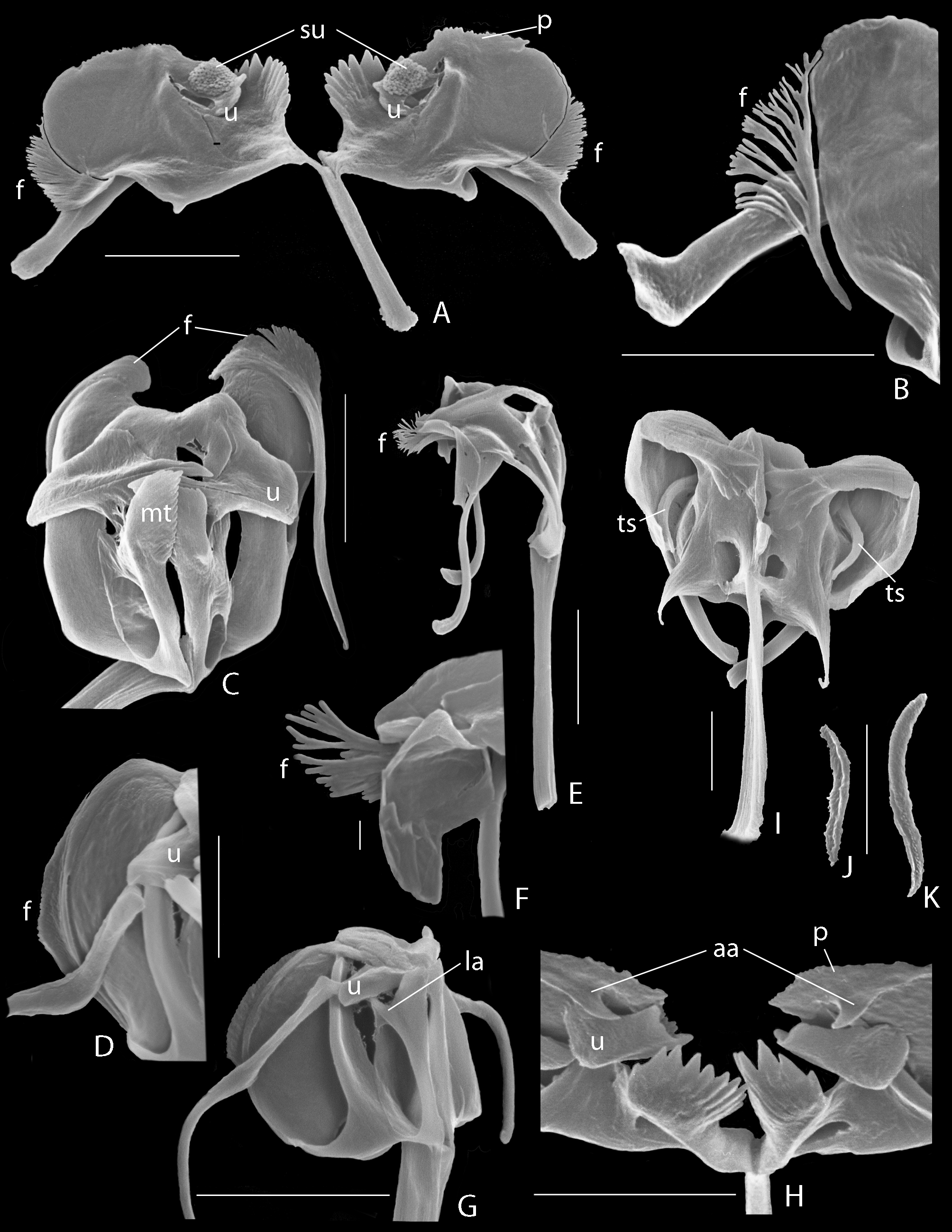
Trophi ( Figs 1 View FIGURE 1 F −H, 2) virgate, moderately asymmetrical; type B according Wulfert (1937, 1938). Rami composed of subbasal, basal and dorsal chambers. Subbasal rami chambers ( Fig. 2 View FIGURE 2 G: sc) elongate, fairly narrow with moderately large, rounded-triangular ventral fenestra proximally ( Fig. 2 View FIGURE 2 G: sf), and comb of 4 strong (left) to 5 smaller (right) strong distal teeth (Fig. I: dt); subdistal lateral apophysis absent. Basal rami chambers ( Fig. 2 View FIGURE 2 G: bc) very shallow with large ventro-lateral fenestra; dorsal margin of chambers extending into fairly acute and long, posteriorly projecting alulae of almost equal length. Dorsal chambers ( Fig. 2 View FIGURE 2 G: dc) very shallow with very large lateral fenestra, anteriorly with distal comb of teeth, the pecten rami ( Figs 2 View FIGURE 2 H, I: p); pectines more or less oriented in the dorso-ventral plane, left one ( Figs 2 View FIGURE 2 K, L) with 7 large appressed distal teeth and ~ 4−5 less obvious very short appressed proximal knobs, right one ( Figs 2 View FIGURE 2 M, N) with 9 large appressed distal teeth and ~ 15−20 short appressed proximal knobs; distal teeth with seam at outer margin; most distal (ventral) teeth largest; apical rami apophyses absent. Flabella rami ( Figs 2 View FIGURE 2 F, G: f; J) fairly small, with reduced fan and shaft, the composing sclerite elements fused. Fulcrum long, in dorsal view slightly expanding towards distal end; in lateral view parallel-sided the greatest part of its length, and slightly expanding proximally and distally; basal fulcrum apophysis absent. Unci with single, acute tooth, and dorsal broadly expanded shaft ( Figs 2 View FIGURE 2 G, I: u); tooth shaft-length; unci with small ventral apophysis near mid-length, dorsally and opposite of it a small apophysis incorporated in expansion of shaft; left tooth with blunt inner lamellar expansion ( Fig. 2 View FIGURE 2 L: u), right tooth ( Figs 2 View FIGURE 2 M, N: u) with 2 large and at least 2 small acute subunci ( Figs 2 View FIGURE 2 M, N: su). Manubria straight, with characteristic asymmetrical distal crutches. Crutch of left manubrium ( Fig. 2 View FIGURE 2 D) sickle-shaped, near its mid-length transversely placed on manubrial shaft; concave side of sickle towards trophi axis; ventral half of sickle widest; outer margin of sickle with short, acute lateral projection. Crutch of right manubrium ( Fig. 2 View FIGURE 2 E) smaller, more or less crooked, ventral half weakly curved, dorsal half weakly expanded dorso-ventrally; outer margin without acute projection. Dorsal and ventral manubrial chambers lamellar, extending till mid-length of shaft; ventral ones distally rounded, dorsal ones terminating into acute, caudally directed and weakly reinforced projection, with lateral opening.
Measurements. Body (N=5): total length 156−170 µm, toe 22−24 µm; trophi (N=4) 27.7−28.6 µm, ramus (incl. alula) 12.2−14.6 µm, fulcrum 18.4−19.4 µm, uncus (l & r) 8.6−8.9 & 6.4−7.0 µm, manubrium (l & r) ~14.2−16.2 & 14.5−15.8 µm, flabellum ~ 3.5 × 0.9 µm.
Distribution and ecology. Cephalodella jersabeki n. sp. is to date only known from its type locality as epibiont of G. pulex .
Comments. The overall more or less similar habitus suggests that Cephalodella jersabeki n. sp. may be related to C. jakubskii Wiszniewski described from Pínsk, Byelorussia, and likewise living in association with G. p u l e x ( Wiszniewski 1953). It is easily distinguished from the latter by the characteristic shape of the crutches of the manubria, especially the left sickle-shaped one with lateral spine (even obviously recognizable by transparency using light microscopy at lower magnification), which are simply T-shaped in C. jakubskii . Other differences concern the toes which are slightly recurved dorsally (straight in C. jakubskii ), and the presence of a tail (lacking in C. jakubskii ). Another species, C. donneri Hauer , described as epibiont of the crayfish Potamobius torrentium Schrank ( Hauer 1959) , was treated as possibly synonymous with C. crassipes (Lord) by Koste (1978) and Nogrady et al. (1995), and considered a junior subjective synonym of C. jakubskii by Jersabek (2002), Jersabek & Leitner (2013) and Jersabek et al. (2015). However, if the descriptions and figures of Wiszniewski (1953) and Hauer (1959) are reliable, C. donneri might be a valid species adapted to another basibiont. In C. jakubskii the manubrial crutches are T-shaped, the left smaller than the right one and without spine, whereas in C. donneri the ventral branches of the crutch are apparently a caudal continuation of the manubrial shaft, and the dorsal branches are pointing antero-dorsally with the left branch bearing a terminal spine; the unci are apparently much longer in C. donneri ; the toes are conical in both species, straight in C. jakubskii and, similar to C. jersabeki n. sp., slightly dorsally recurved distally in C. donneri .
In the most recent revision of the genus Cephalodella by Nogrady et al. (1995), C. jersabeki n. sp. keys out to C. crassipes , but the trophi type of the latter was erroneously called type B in the key, whereas it was described as type A in the species description. They also considered C. jakubskii and C. donneri as possible synonyms of C. crassipes , notwithstanding the significant differences of their diagnostic features (for a discussion of the nomenclatorial confusion concerning these species see Jersabek (2002)). Inclusion of C. jersabeki n. sp. in the C. crassipes complex, as Jersabek (2002) did for C. jakubskii , is not followed here in view of the obvious differences in trophi structure, i.e. the presence of crutched manubrial cauda and the pronounced alulae on the rami which are both lacking in C. crassipes , and the unambiguously differently shaped rami teeth which are distal instead of medial as in the latter.
The study by light microscopy of the rami structure in Cephalodella is difficult and even in the few studies by SEM there is some disagreement in the interpretation and naming of the different composing structures ( Markevich 1989; Segers & Pholpunthin 1997; De Smet 1998a; Segers & Chittapun 2001; Sørensen 2002; Jersabek et al. 2011; Fischer & Ahlrichs 2006, 2011). In the present paper we recognize three chambers composing the rami ( Fig. 2 View FIGURE 2 G): subbasal, basal and dorsal, which can be traced in all species studied to date (unpubl. res.). The subbasal chambers (basal chambers in Markevich (1989), De Smet (1998a); basal apophyses in Segers & Pholpunthin (1997), Segers & Chittapun (2001); subbasal chambers in Sørensen (2002: Fig. 6 View FIGURE 6 ), Jersabek et al. (2011), Fischer & Ahlrichs (2006, 2011)) are the much isolated, often long and narrow ventral chambers ( Fig. 2 View FIGURE 2 G: sc) with proximal ventral or ventro-lateral basifenestra and distal ( Fig. 2 View FIGURE 2 I: dt) or medial ( Fig. 4 View FIGURE 4 C: mt) teeth. The basal chambers (subbasal in Markevich (1989), De Smet (1998a); basal in Jersabek et al. (2011), Fischer & Ahlrichs (2011); rami in Segers & Pholpunthin (1997), Fischer & Ahlrichs (2006)) form the medio-ventral section of the widely arched part of the rami ( Fig. 2 View FIGURE 2 G: bc). They are elongate and more or less rounded-triangular and fairly shallow to deep chambers with very large opening, separated from the dorsal chambers by a pronounced rim running from the base of the fulcrum or caudal ramus margin towards the dorso-apical ramus margin. This rim is also easily recognized by LM and shown in most of the pictures obtained by the latter research method (e.g. Wulfert 1937, 1938; Nogrady et al. 1995). The dorsal chambers ( Fig. 2 View FIGURE 2 G: dc) form the much arched plate-shaped part of the rami with dorsally rounded margin and very large shallow to moderately deep fenestra.
The term pecten rami (Latin pecten, comb) is introduced to denote the comb of appressed projections at the medio-frontal margin of the dorsal rami chambers ( Figs 2 View FIGURE 2 , 3 View FIGURE 3 : p). Three types of rami pecten may be grossly distinguished. The combs can be composed of a fairly large series of similar delicate projections ( Figs 3 View FIGURE 3 A, B: p), particularly present in Cephalodella having type A trophi, e.g. C. hoodii (Gosse) , C. delicata Wulfert , C. ventripes (Dixon-Nutall) and C. acidophila Jersabek, Weithoff & Weisse (Jersabek et al. 2011 ; Jersabek, pers. comm.), but may also be present in species of trophi type C such as C. maior (Zavadovski) ( Fig. 3 View FIGURE 3 C: p). Fusion of the projections results in a medially more or less strongly crenate plate, such as in C. panarista Myers ( Fig. 3 View FIGURE 3 E: p).The ramus pecten can also be shaped as a comb of stout teeth of more or less similar shape ( Fig. 3 View FIGURE 3 F: p, C. stenroosi Wulfert ), or as a differentiated comb with a stout major ventral tooth, followed by a series of smaller stout teeth, and terminating in a series of short dorsal knobs such as in C. jersabeki n. sp. ( Figs 2 View FIGURE 2 H, I, K −N) and C. boettgeri Koste ( Fig. 3 View FIGURE 3 D: p). Teeth of this third pecten type are provided with a longitudinal seam at their outer surface. Species with plate-shaped or toothed pectines are most common in species of trophi types B and C.
The term apical ramus apophysis is proposed for the protuberance at the outer surface/margin of the dorsal rami chamber posterior or off the pecten. Depending on the species the apical apophyses can be absent (e.g. C. jersabeki n. sp.) or present. If present, they may be thorn-shaped such as in C. theodora Koch-Althaus ( Fig. 4 View FIGURE 4 H: aa), tooth-like as in C. edax Hollowday ( Jersabek 2003, Fig. 2 View FIGURE 2 ), or irregularly shaped as in C. boettgeri ( Fig. 3 View FIGURE 3 D: aa).
The term subdistal lateral ramus apophysis is proposed for the lateral apophysis posterior from the distal comb of teeth of the subbasal chambers ( Fig. 4 View FIGURE 4 G: la), and apparently connected with the unci.
The term flabellum (diminutive of Latin flabrum, fan) rami is introduced for the fan-shaped elements associated with the dorsal margin of the dorsal rami chambers ( Figs 4 View FIGURE 4 A −E). The flabella are basically composed of a series of elongate sclerite elements forming the rays of the fan. The sclerite elements are provided with a seam, and grouped into a more or less developed stalk caudally, and more or less diverging into a unilaterally fanned structure antero-dorsally. Their ventral margin is closely connected to the dorsal margin of the dorsal chambers, but in species showing flabella with long stalk, the stalk may become more or less free from the rami ( Fig. 4 View FIGURE 4 C: f). The rays of the fan may be simple, bifid at their free end, or more or less strongly branching ( Fig. 4 View FIGURE 4 B: f); they may be spread ( Fig. 4 View FIGURE 4 B: f) or compacted into a broom-like structure ( Fig. 4 View FIGURE 4 C: f). In species with reduced flabellum the composing sclerite elements are usually fused into a platelet ( Fig. 2 View FIGURE 2 J).
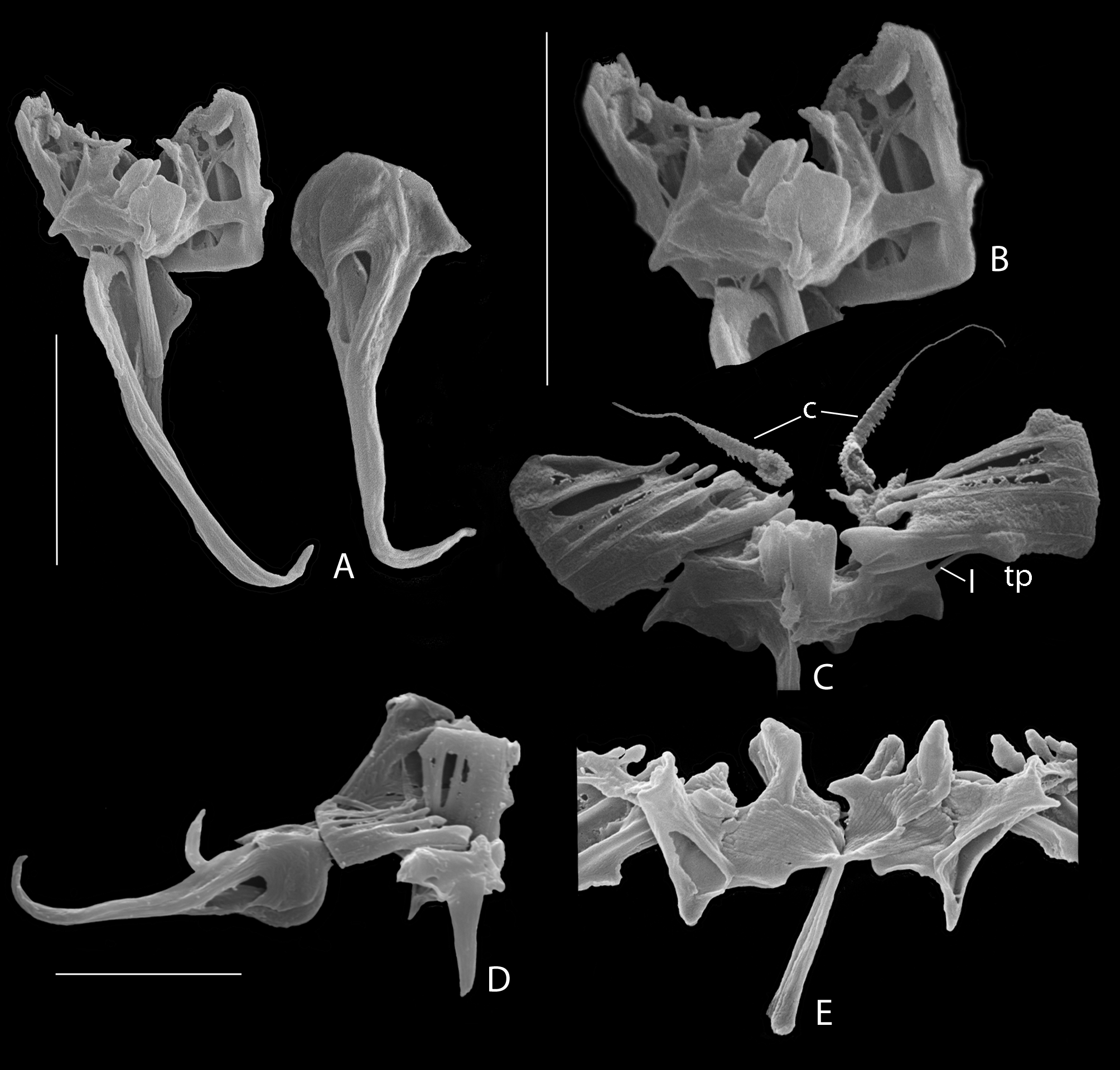
Apparently, flabella rami are present only in the genus Cephalodella , and have been called subunci (e.g. Wulfert 1937; Donner 1950; Koste 1978; Nogrady et al. 1995; Fischer & Ahlrichs 2011), or pleural rods (e.g. De Smet 2001; Sørensen 2002, Fig. 6 View FIGURE 6 ). The latter terms are also used to denote other, different and unrelated structures. The Figs 4 View FIGURE 4 A −G clearly show that the flabella are not associated with the unci, and the term subunci should only be used for the hard structures situated distally at the inner side of the unci ( Figs 2 View FIGURE 2 M, N; 4A: su). The term pleural rod has been used for the long, rod-shaped straight (in numbers of one, 2 or 3), U, V, or Y-shaped structures found laterally to medio-ventrally from the incus and more or less in line with the trophi axis, and hitherto only demonstrated in the genus Cephalodella . They easily dissolve in hypochlorite solution, suggesting that they may belong to the ligamentous system of the trophi or supporting structure of the mastax. The term pleural rod has been used in other Notommatidae as well (e.g. Eothinia , Notommata , Resticula ) to denote the paired and strongly sclerified, straight or slightly bent rod-shaped trophi elements ( Figs 4 View FIGURE 4 I: ts, 4J, K), often with small terminal widening dorsally, situated between the head of the manubria and the rami, and more or less perpendicular to the trophi axis. We propose to keep ‘pleural rod’ for the long, longitudinally oriented and easily dissolving structures in Cephalodella , and to use ‘transversal sclerite’ for the elements located transversally between malleus and incus in the other notommatid genera.
Fischer & Ahlrichs (2011) introduced a new definition scheme for the trophi of Cephalodella based on SEM to identify the six types originally described by Wulfert (1937, 1938). However, this new scheme, based on trophi preparations obtained with the dithio-dl-threitol method modified after Kleinow et al. (1990), can be confusing as some delicate trophi parts may be missed by this method. For example, they do not mention flabella in their group with trophi types A, E and F, but use this feature to differentiate these types with trophi type D species, notwithstanding that these structures are clearly seen in preparations obtained by the NaOCl method (e.g. C. hoodii ( Fig. 3 View FIGURE 3 A: f) and C. megalocephala (Glascott) , ( Figs 4 View FIGURE 4 E, F: f), respectively type E and F according Fischer & Ahlrichs l.c.).


| RBINS |
Royal Belgian Institute of Natural Sciences |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Monogononta |
|
Order |
|
|
Family |
|
|
Genus |