
Halichoeres rubrovirens, Rocha & Pinheiro & Gasparini, 2010
|
publication ID |
https://doi.org/ 10.11646/zootaxa.2422.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:C11A5CD4-1794-4D05-B48A-FA1A06E3EBC2 |
|
persistent identifier |
https://treatment.plazi.org/id/7D1E4CCC-FE91-4BFC-A5F9-F86E45AABBE8 |
|
taxon LSID |
lsid:zoobank.org:act:7D1E4CCC-FE91-4BFC-A5F9-F86E45AABBE8 |
|
treatment provided by |
Felipe |
|
scientific name |
Halichoeres rubrovirens |
| status |
sp. nov. |
Halichoeres rubrovirens View in CoL sp. n.
Figures 1–5 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 , Table 1
Holotype. CIUFES 1470, adult, 231 mm, Trindade Island , Brazil, Calhetas Reef, 20º30’18’’S, 29º18’30’’W, 12 m, spear gun, H. Pinheiro, 24 April 2007. GoogleMaps
Paratypes. CIUFES 1471, adult, 219 mm, same locality as holotype, 15 m, spear gun, H. T. Pinheiro, 29 April 2009 GoogleMaps . CIUFES 1472, adult, 173 mm, same locality as holotype, 15 m, spear gun, H. T. Pinheiro, 01 May 2009 GoogleMaps . CIUFES 1473, subadult, 107 mm, same locality as holotype, 15 m, hand nets, H. T. Pinheiro, 29 April 2009 GoogleMaps . CIUFES 0317, juvenile, 54.3 mm, same locality as holotype, 12 m, hand nets, H. T. Pinheiro, 18 April 2007 GoogleMaps . CIUFES 1279, juvenile, 48.3 mm, same locality as holotype, 12 m, hand nets, H. T. Pinheiro, 24 April 2007 GoogleMaps . CIUFES 1474, juvenile, 60.4 mm, same locality as holotype, 12 m, hand nets, H. T. Pinheiro, 24 April 2007 GoogleMaps . CIUFES 1475, juvenile, 57.8 mm, same locality as holotype, 12 m, hand nets, H. T. Pinheiro, 24 April 2007 GoogleMaps . CIUFES 1476, juvenile, 72.8 mm, same locality as holotype, 12 m, hand nets, H. T. Pinheiro, 24 April 2007 GoogleMaps . MZUSP 104530 View Materials (HTP 667 “C”), adult, 206.0 mm, same locality as holotype, 15 m, spear gun, H. T. Pinheiro, 29 April 2009 GoogleMaps . UF 175341 (HTP 669 “E”), adult, 168.0 mm, same locality as holotype, 15 m, spear gun, H. T. Pinheiro, 29 April 2009 GoogleMaps . USNM 397005 About USNM (HTP 786 “G”), juvenile, 79.0 mm, same locality as holotype, 15 m, hand nets, H. T. Pinheiro, 20 April 2009 GoogleMaps .
Diagnosis. Halichoeres rubrovirens differs from all of its New World congeners by the following combination of characters: dorsal rays IX, 11 (one individual with IX, 10); anal rays III, 12; pectoral rays 13; lateral-line scales 27 (one individual with 25); no scales on cheek or opercle; gill rakers 12–17 (modally 14); body depth 4.6–5.1 in SL; pair of forward-projecting, slightly outcurved, canine teeth at front of upper jaw, two pairs in lower jaw, outer pair also curving laterally; color in life of adult yellowish-green, the body with two broad dotted orange-pink stripes, the 1st running along the dorsum from above the eye to the upper caudal peduncle; 2nd stripe from upper opercle margin to base of caudal fin; series of irregular orange-pink spots along the lateral-line; bright blue spot on upper pectoral-fin base; upper third margin of pectoral-fin dark.
Description. Dorsal rays IX, 11 (one individual with IX, 10); anal rays III, 12; all dorsal and anal rays branched, the last to base; pectoral rays 13, the uppermost unbranched; pelvic rays I, 5; principal caudal rays 18, the upper and lower unbranched; lateral-line scales 27 (one individual with 25); two to three scale rows above lateral line to origin and middle of dorsal fin; eight scale rows below lateral line to origin of anal fin; circumpeduncular scales 13–14; gill rakers 17 (12–17, modally 14); vertebrae 24.
Body slender, depth 3.36 (3.29–5.08) in SL, and moderately compressed, width 2.46 (1.66–2.46) in body depth; head length 4.2 (3.30–4.34) in SL; snout length 2.35 (2.35–2.67 in adults; 2.93–3.65 in juveniles); orbit diameter 6.83 (5.58–6.83 in adults; 3.85–4.84 in juveniles) in head length; caudal peduncle depth 1.96 (1.83– 1.96 in adults; 2.07–3.12 in juveniles) in head length; caudal peduncle length 2.52 (2.44–3.21 in adults; 1.74– 2.33 in juveniles) in head length; opercular flap extending slightly beyond the upper end of pectoral fin base but not beyond the lower end of pectoral fin base; dorsal profile of head uniformly convex; eye above the midlateral line of head, distance above eye to dorsal edge of head nearly half than distance below eye to ventral edge.
Mouth terminal, protusible, and slightly oblique, upper jaw forming angle of about 30° to horizontal axis of head and body; snout moderately pointed, its length 2.35 (2.35–3.5) in head length; pair of forwardprojecting, slightly outcurved, canine teeth at front of upper jaw, two pairs in lower jaw, outer pair also curving laterally; posterior canine sharp and well developed.
Anterior nostril a very small membranous tube one-half distance on a line from top of fleshy edge of orbit to median base of groove behind upper lip; posterior nostril dorsoposterior to anterior nostril, larger than sensory pores of head, but largely covered by anterior flap; internarial distance two-thirds pupil diameter; sensory pores (double pores counted as one) in series beginning on front of snout, passing above nostrils, and encircling orbit 19 (19–21); pores of preopercular-mandibular series 16 (15–16).
Scales cycloid; lateral line continuous, deflected downward below last three rays of dorsal fin, extending to a straight peduncular part; anterior lateral-line scales with three pores, posterior scales with single pore; nostrils with anterior short tube and posterior flap; caudal fin truncate to slightly rounded; free vertical and horizontal margins of preopercle smooth; scales end above preopercle on nape, head naked; scales on chest smaller than body, extending anteriorly to rear margin of isthmus; single large, elongate scale between pelvicfin bases; four rows of scales on caudal fin, gradually decreasing in size to rear of lateral line.
Origin of dorsal fin above 2nd lateral-line scale, the predorsal length 4.13 (4.06–4.28 in adults; 3.14–3.78 in juveniles) in SL; dorsal spines progressively longer, the 1st 4.95 (4.28–6.02) in head length, and the longest (10th or 11th) 2.26 (2.13–2.78) in head length; origin of anal fin below base of 1st dorsal soft ray, the preanal length 1.8 (1.75–1.8) in SL; 1st anal spine very slender, 6.2 (5.7–6.65) in head length; 3rd anal spine 3.4 (3.35-3.6) in head length; penultimate anal soft ray longest, 2.l (1.9–2.1) in head length; caudal fin rounded, 1.25 (1.25–1.35) in head length, 4.05 (4.05–4.3) in SL; 3rd pectoral ray longest,1.45 (1.4–1.55) in head length; pelvic fins nearly or just reaching anus in adult males, 1.25 (1.25–1.3) in head length; in females, 1.5– 1.55 in head length.
Color of holotype in alcohol. Body light brown ventrally, darker dorsally; head light yellowish brown, lighter ventrally, with dark brown area below and in front of eye; a dark stripe radiating backwards from eye to opercle, bifurcating and turning down towards pectoral fin base; fins translucent pale yellowish, the upper posterior corner of pectoral fin with an indistinct dark spot; caudal fin with dark posterior margin ( Fig. 1 View FIGURE 1 ).
Color in life. Color of live adults as in Figure 2 View FIGURE 2 . Figures 3A, and 3B View FIGURE 3 are of the intermediate and juvenile color phases.
Comparisons to congeners. The striking red/green adult color pattern readily distinguishes H. rubrovirens from all of its congeners in the New World. Among its Atlantic congeners, H. rubrovirens is most similar to H. bivittatus , with whom it shares most counts and proportions. Juveniles are similar in color to H. bivittatus ( H. bivittatus also has two dark stripes along the body, but the ventral stripe is less intense than the dorsal stripe, whereas the two stripes are equally intense in H. rubrovirens ), however, H. rubrovirens has modally less gill rakers than H. bivittatus (14 in H. rubrovirens versus 17 in H. bivittatus ).
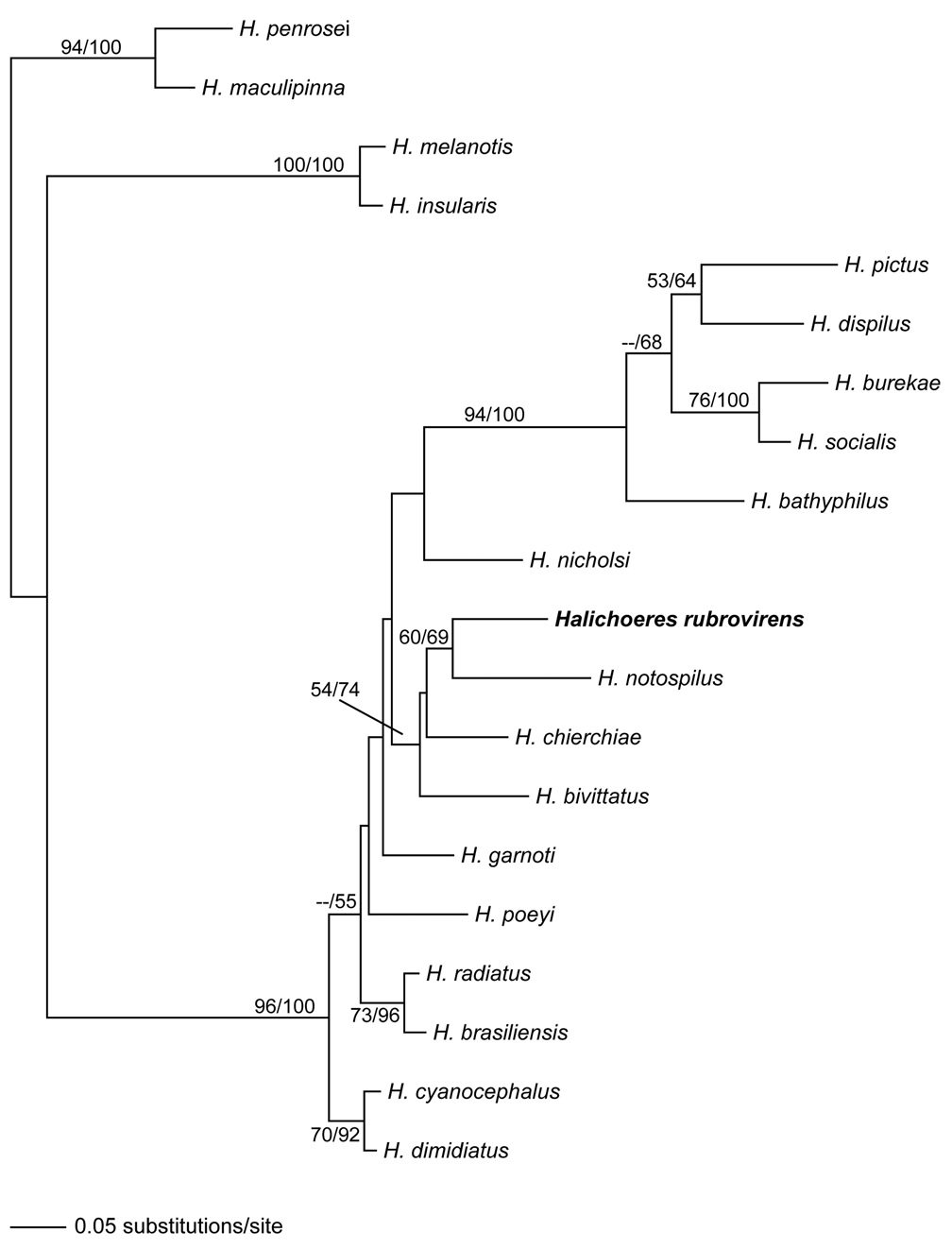
In addition to H. bivittatus , H. rubrovirens also shares most of its counts with two eastern Pacific species, H. chierchiae and H. notospilus . These four species form a well-supported monophyletic clade (the “ bivittatus ” clade) in our mitochondrial DNA phylogeny ( Fig. 4 View FIGURE 4 ). They share the same counts for dorsal, pectoral and anal fin rays, very similar counts for lateral-line scales and gill rakers, have very similar morphometric measurements and somewhat similar color patterns (stripes of varying intensity can be observed in all four species at least during one stage of development), but can be easily separated based on color in all stages of life. H. garnoti and H. poeyi are the only two other Atlantic species with which H. rubrovirens shares its number of dorsal (IX, 11), pectoral (13) and anal fin rays (III, 12), however, these can be readily distinguished from the new species by color. Even though there is some overlap in gill raker counts, this measure can also distinguish H. rubrovirens from H. garnoti and H. poeyi (modally 14 in H. rubrovirens , 17 in H. garnoti and 18 in H. poeyi ) Not surprisingly, H. garnoti and H. poeyi seem to be in an unresolved position on our phylogenetic analysis in relation to the bivittatus clade ( Fig. 4 View FIGURE 4 ).
Genetics. We obtained partial (515 base pairs long) CytB sequences from paratypes of Halichoeres rubrovirens and an additional 19 species of New World Halichoeres CytB sequences available in Genbank (some for the fist time in this study, others from Rocha 2004; Rocha et al. 2005; Weaver & Rocha 2007; Luiz Jr et al. 2009) to estimate the phylogenetic position of the new species. The resulting Bayesian phylogenetic tree ( Fig. 4 View FIGURE 4 ) shows (as does its distinctive color pattern) that H. rubrovirens is clearly a new species, well separated from all of its congeners.
The most surprising result from the genetic analysis is that it shows a tropical eastern Pacific species, H. notospilus , as the sister species to H. rubrovirens . This result indicates that today’s population of H. rubrovirens is probably a relict of a species with a much wider distribution, spamming the entire tropical western Atlantic. The genetic divergence between H. rubrovirens and H. notospilus further supports this hypothesis. It is virtually the same as the one between the pair H. pictus and H. dispilus , which is considered a trans-Isthmian species pair separated by the final closure of the Isthmus of Panama ( Barber & Bellwood 2005). Alternatively, speciation events that separated the four species in the bivittatus clade may have occurred during a short time spam, and H. rubrovirens may be a distant sister to H. bivittatus . However, our hypothesis is based only on mtDNA, and relationships among species in this group may change with the addition of nuclear loci data as exemplified by grunts, a group in which several discordances were found between nuclear and mitochondrial DNA datasets ( Rocha et al. 2008).
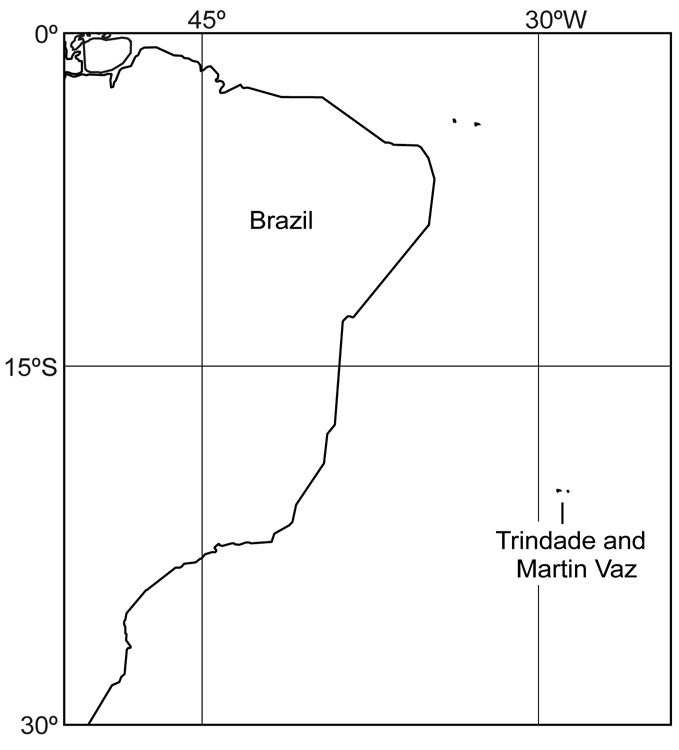
Distribution and Ecology. Halichoeres rubrovirens is endemic to the Trindade and Martin Vaz Island group, located 1200 km east of the southeastern Brazilian coast ( Fig. 5 View FIGURE 5 ). Little is known about its ecology, but juveniles seem to mimic and usually swim together with schools of Thalassoma noronhanum , which they resemble in color. Specimens were observed over high rugosity rocky bottoms with calcareous algae at depths between 6 and 25 m.
Etymology. We have selected the name rubrovirens from the Latin, which means red and green. The name is an allusion to the distinctive red and green color pattern of the adults.



| T |
Tavera, Department of Geology and Geophysics |
| UF |
Florida Museum of Natural History- Zoology, Paleontology and Paleobotany |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |