
Pristimantis boulengeri
|
publication ID |
https://doi.org/10.11646/zootaxa.4243.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:B4D35FB5-BC82-426C-BFCF-4EEF4D6EAFE6 |
|
DOI |
https://doi.org/10.5281/zenodo.5631800 |
|
persistent identifier |
https://treatment.plazi.org/id/EE681907-A041-FFE3-FF68-C7E3FDDEBDDB |
|
treatment provided by |
Plazi |
|
scientific name |
Pristimantis boulengeri |
| status |
|
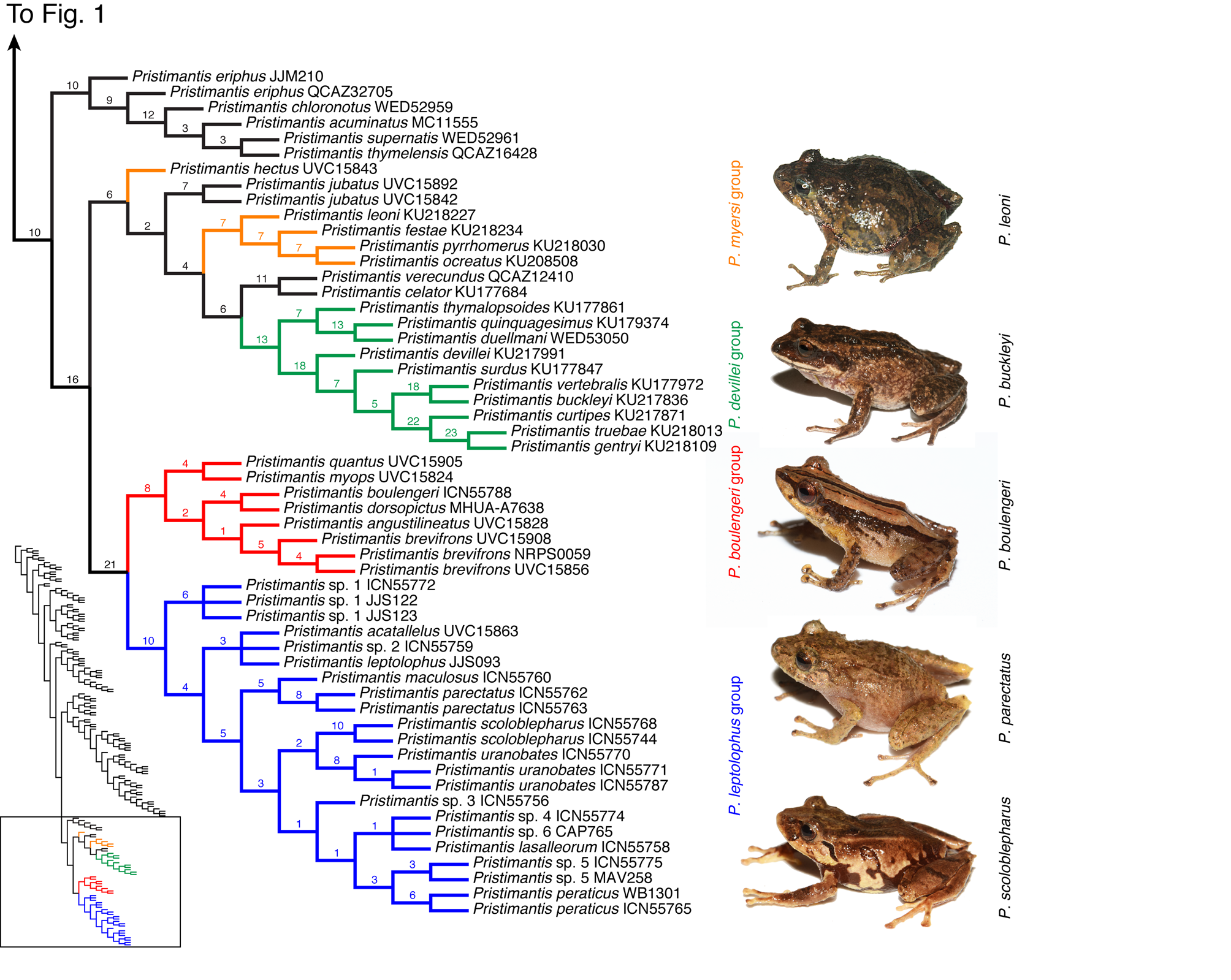
Pristimantis boulengeri species group
Content ( Fig. 2 View FIGURE 2 ). Eight recognized species: Pristimantis angustilineatus (Lynch, 1998) , P. baiotis (Lynch, 1998) , P. boulengeri ( Lynch, 1981) , P. brevifrons ( Lynch, 1981) , P. dorsopictus ( Rivero & Serna, 1988) , P. myops (Lynch, 1998) , P. quantus (Lynch, 1998) , and P. urani ( Rivera-Correa & Daza, 2016) .
Distribution. The species are known from the Cordillera Occidental of Colombia (from Munchique National Park to north of the Orquideas National Park, at elevations of 1780–2610 m) and along the Cordillera Central of Colombia (from Cauca Department to Antioquia Department, 2500–3200 m).
Morphological diagnosis. Males with vocal slits and external vocal sac. Dorsolateral folds absent (with exception of Pristimantis quantus ). Ventral skin coarsely areolate. Parietal peritoneum covered by iridophores (condition unknown in P. quantus and P. myops ). Nuptial pads present on Finger I (absent in P. myops ). Ulnar tubercles present (with exception of P. brevifrons ). Tip of snout with small papilla. Double distal subarticular tubercle on Finger III (polymorphic in P. brevifrons ). Double distal subarticular tubercle present on Finger IV (polymorphic in P. brevifrons and P. boulengeri ). Heel tubercle present (with exception of P. angustilineatus ). Finger I shorter than Finger II. Digital discs expanded and round, except in Finger I and Toe I in P. myops , P. quantus and P. brevifrons . Toe V much longer than Toe III, reaching level of distal subarticular tubercle of Toe IV. Cranial crests absent. SQp portion of m. depressor mandibulae reaching dorsally to level of optic ramus of squamosal.
We propose as putative synapomorphies 1) presence of a double distal subarticular tubercle on Finger III, 2) presence of a double distal subarticular tubercle on Finger IV, 3) parietal peritoneum covered by iridophores (unknown in P. myops and P. quantus ), 4) extended external vocal sac; and 5) tip of the snout with small papilla.
Comments. Pristimantis brevifrons , P. boulengeri and P. dorsopictus were species of the P. lacrimosus group, which is not monophyletic according to our results. Hedges et al. (2008) included three of the 18 species then placed in the P. lacrimosus group, and those three species also form a clade in our results. Hedges et al. (2008) characterized the group as arboreal and commonly inhabiting bromeliads. Pinto-Sánchez et al. (2012) found the species group to be paraphyletic, with P. brevifrons falling outside the clade. Padial et al. (2014a) also failed to corroborate the monophyly of the P. lacrimosus group due to the placement of P. acuminatus , a species of P. unistrigatus species group sensu Hedges et al. (2008), inside the group. They transferred this and several other species to the P. lacrimosus group, due to the presence of an acuminate snout, smooth dorsal skin, and round and ovate finger and toe discs, bringing the total number of species to 25. Mendoza et al. (2015) sampled more specimens of P. brevifrons and also recovered this species outside the P. lacrimosus species group, but also found P. moro to be placed inside the group. Rivera-Prieto et al. (2015) included P. dorsopictus in their analysis and found it to be placed outside the P. lacrimosus species group. Heinicke et al. (2015) sampled P. prolixodiscus , then a member of the P. lacrimosus species group and, finding that species to fall outside Pristimantis , designated it as the type species of their new genus Tachiramantis . Rivera-Correa and Daza (2016) found P. lacrimosus group to be paraphyletic, noting that the species formed two clades (their clades A and B). They also describe a new species, P. urani , as the sister of P. brevifrons according to their phylogenetic hypothesis.
DNA sequences are unavailable for Pristimantis lacrimosus and its phylogenetic position has never been tested. Lynch and Schwartz (1971) redescribed P. lacrimosus , and Heyer and Hardy (1991) designated the specimen figured by Lynch and Schwartz (1971) as neotype. Neither the figure nor the redescription show double distal subarticular tubercles on Fingers III or IV, putative synapomorphies of the P. boulengeri group, proposed herein. On that basis, we hypothesize that P. lacrimosus is not part of this clade, which is the reason we propose the P. boulengeri group as a new group. DNA sequences are also unavailable for Pristimantis baiotis and its phylogenetic position also has not been tested, so it is here tentatively allocated based on putative morphological synapomorphies, especially the double subarticular tubercles (Lynch, 1998). The description of P. urani was published ( Rivera and Daza, 2016) after analyses for the present study had been completed. As such, we include it in the P. boulengeri group on the basis of Rivera and Daza’s (2016) phylogenetic analysis.
The Pristimantis boulengeri and P. leptolophus species groups form a monophyletic group with GB = 21. Pristimantis brevifrons , P. angustilineatus , P. boulengeri , and P. dorsopictus can inhabit bromeliads ( Lynch, 1981; Lynch, 1998; Rivero and Serna, 1988).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Brachycephaloidea |
|
Family |
|
|
Genus |