
Baeolidia moebii Bergh, 1888
|
publication ID |
https://doi.org/10.11646/zootaxa.3802.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:4095CA09-8EA4-4941-8286-32E95F0206AE |
|
DOI |
https://doi.org/10.5281/zenodo.6134190 |
|
persistent identifier |
https://treatment.plazi.org/id/ED36FA51-A03E-FF84-FF1B-FC35E19EF886 |
|
treatment provided by |
Plazi |
|
scientific name |
Baeolidia moebii Bergh, 1888 |
| status |
|
Baeolidia moebii Bergh, 1888 View in CoL
( Figs. 2 View FIGURE 2 A–B, 3A, 4A–B, 5, 6A)
Baeolidia moebii Bergh, 1888: 778 View in CoL , pl. LXXIX, Figs. 10–16 View FIGURE 10 View FIGURE 11 View FIGURE 12 View FIGURE 13 View FIGURE 14 View FIGURE 15 View FIGURE 16 , pl. LXXX, Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 . Baeolidia major Eliot, 1903: 252 View in CoL .
Baeolidia major amakusana Baba, 1937: 335 View in CoL .
Berghia major ( Eliot, 1903) View in CoL : Edmunds 1969, 467, Fig. 10 View FIGURE 10 . Spurilla major ( Eliot, 1903) View in CoL : Rudman 1982, 169, Figs. 18 View FIGURE 18 –19.
Type locality. Grand Bay, Rodrigues Island, Mauritius.
Type material. To our knowledge no type material remains. We designate the specimen MNCN 15.05/63447 from Mozambique as the neotype in order to avoid confusion with similar species.
Material examined. Neotype: MNCN 15.05/63447, one specimen, dissected, 18 mm in length preserved, Mozambique, Zavora Bay, collected by Yara Tibiriça, January 2012. Other material: CASIZ 177602, one specimen, dissected, 10 mm in length preserved, Philippines, Luzon Island, Batangas Bay, Calumpan Peninsula, collected by Terrence M. Gosliner, 16 April 2008; CASIZ 157017, two specimens, dissected, 25 and 24 mm in length preserved, Philippines, Luzon Island, Batangas Province, Balayan Bay, Seafari Beach, collected by Ángel Valdés and Mary James Adams, 12 May 2001; CASIZ 186211, one specimen, dissected, 10 mm in length preserved, Philippines, Luzon, Batangas Province, Calumpan Peninsula, Batangas Bay, collected by Alicia Hermosillo, 0 4 May 2011; CASIZ 180327, one specimen, dissected, 6 mm in length preserved, Hawaii, Maui, Airport Beach, collected by Cory Pittman, 0 4 October 2008; CASIZ 0 76055, one specimen, dissected, 16 mm in length preserved, Mexico, Baja California, Gulf of California, San Jose Island, Punta Colorado, collected by Antonio J. Ferreira 15 July 1971.
Geographical distribution. Originally described from Mauritius ( Bergh 1888), it is also known from the Seychelles (Gosliner et al. 2008), Tanzania ( Eliot 1903; Edmunds 1970), Mozambique (present study), the Reunion Island ( Bidgrain 2013), Australia ( Cobb & Mullins 2013), Indonesia (Gosliner et al. 2008), the Philippines (Gosliner et al. 2008), Japan ( Baba 1933b, 1937, 1955; Ono 1999, 2004; Nakano 2004) and the Marshall Islands (present study). This species is also found in the Eastern Pacific: Hawaii ( Kay 1979; Gosliner 1980), tropical Pacific North America (Gosliner et al. 2008) and Mexico ( Farmer 1966). Recently, it has also been reported in Turkey ( Turk & Furlan 2011).
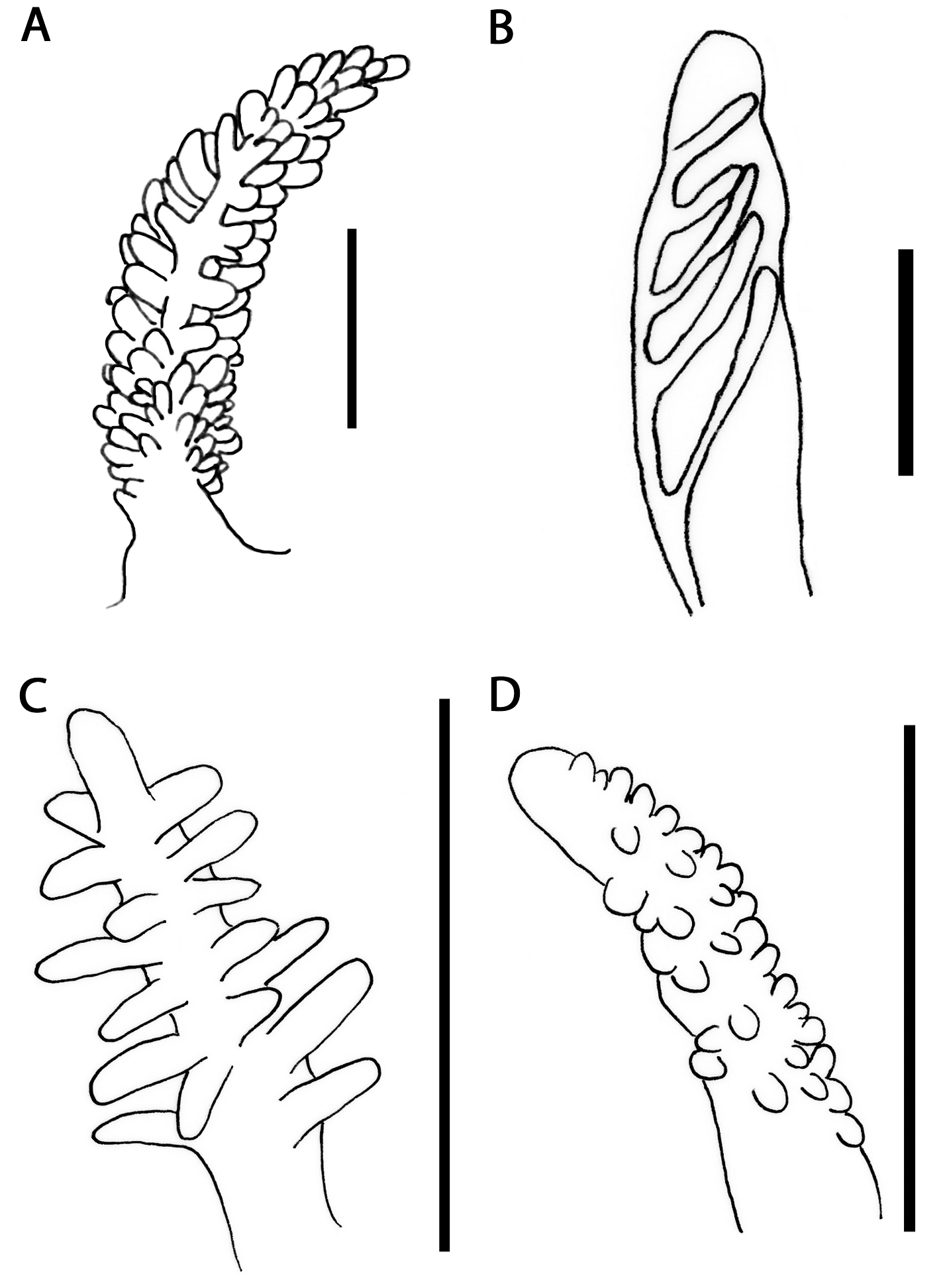
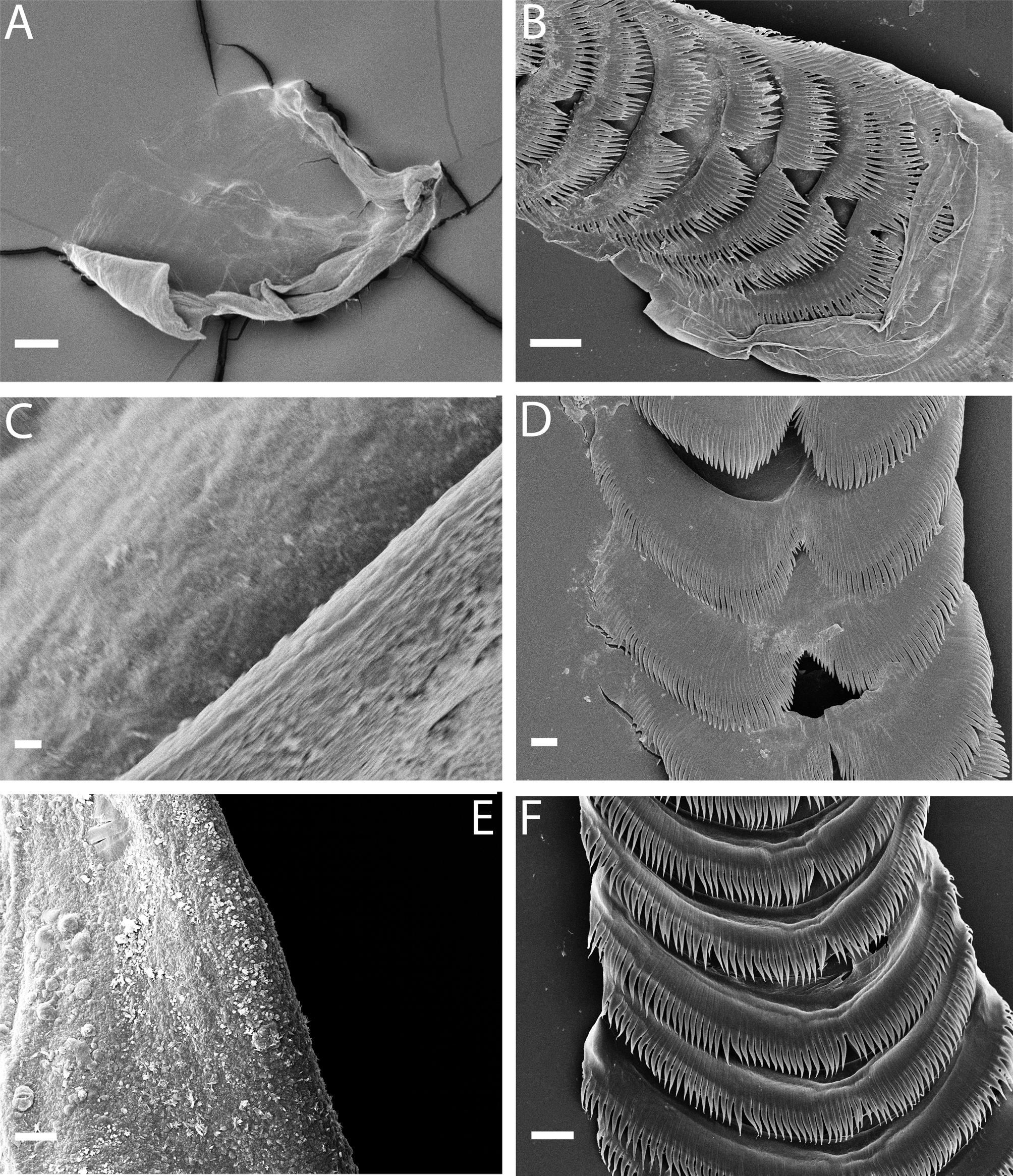
External morphology ( Figs. 2 View FIGURE 2 A–B, 3A): Body large, broad, tapering gradually towards posterior end of foot. Foot corners tentaculiform. Body colour variable, commonly greyish white or brownish green ( Fig. 2 View FIGURE 2 A). Some specimens reddish ( Fig. 2 View FIGURE 2 B). Background colour brownish green, overlaid with reticulate ochre pattern. Dorsum usually with bright white patches. Whitish or yellow ring on anteriormost edge of head, continuing towards oral tentacles and forming a distinct light band; both, ring and band, may be interrupted. Rhinophores, oral tentacles and foot corners same as body colour. Rhinophores shorter than oral tentacles. Rhinophores studded with minute knobs ( Fig. 3 View FIGURE 3 A); apex white. Oral tentacles with white tips.
Cerata length variable. Cerata flattened, almost leaf-like. Branches of digestive gland brownish-green, shining through translucent body wall. Whitish patches on outer side of cerata, probably with bright yellow marks. Cerata usually with bright yellow subapical band and just below it a purplish area. Apex translucent white. Cerata composed of up to nine arches, leaving a distinct gap between pre and post-pericardial groups. Each arch with 4–15 cerata, decreasing in size towards foot. Anus cleioproctic, within second right arch. Genital opening placed among cerata of anteriormost group on right side.
Anatomy. Masticatory border of jaws smooth ( Fig. 4 View FIGURE 4 A). Radular formulae 17 x 0.1.0 ( MNCN 15.05/ 63447, 18 mm), 19 x 0.1.0 ( CASIZ 0 76055, 16 mm), 20 x 0.1.0 ( CASIZ 177602, 10 mm) and 22 x 0.1.0 ( CASIZ 180327, 6 mm). Radular teeth bent upwards with up to 100 elongate, fine and acutely pointed denticles from side to side, without central notch or central cusp ( Fig. 4 View FIGURE 4 B). Teeth progressively smaller towards posterior region of radula. Oral glands fragile, small and spongy, lying dorso-laterally to buccal bulb ( Fig. 5). Oral gland morphology variable, from rounded to elongate. Salivary glands large.
Reproductive system diaulic ( Fig. 6 View FIGURE 6 A). Preampullary duct widening into elongate ampulla. Postampullary duct dividing into oviduct and vas deferens. Vas deferens elongate, moderately wide, penetrating into wider proximal portion of penial sac with unarmed penial papilla. Receptaculum seminis ovoid, short stalk connecting to short oviduct, before latter forms female glands. Vagina ventral to penis.
Remarks. This species has not been studied under the name of Baeolidia moebii since its original description ( Bergh 1888). When Eliot (1903) described B. major , he pointed out that the latter could be merely a full-grown individual of B. moebii since the main difference between both species was the size. Nevertheless, Eliot (1903) finally described B. major as a valid species, owing to the ambiguities in the original description of B. moebii . Taking into consideration Eliot’s statements together with the proximity of both type localities ( Mauritius for B. moebii and Zanzibar for B. major ), Carmona et al. (2013) considered B. major as a junior synonym of B. moebii . Neither Bergh (1888) nor Eliot (1903) mentioned the presence of oral glands in B. moebii . Only Rudman (1982) pointed out the existence of salivary glands in this species. Two of our specimens possessed oral glands, and salivary glands were found twice. This variability could be an artefact of the state of preservation of these specimens. An alternative hypothesis could be an intraspecific variation for these two characters, which has also been observed in Limenandra nodosa ( Carmona et al. 2014c).
Despite the large variability in colouration, our specimens identified as Baeolidia moebii match with the information given by Bergh (1888) and Eliot (1903) as well as with more recent descriptions ( Edmunds 1969; Gosliner 1980; Rudman 1982). Gosliner (1985) stated the presence of a penial gland in the reproductive system of B. moebii (identified as Berghia major ). The only aeolidiid where this condition has been described is Limenandra nodosa ( Schmekel 1970) , although Carmona et al. (2014c) did not find any traces of a penial gland. Since none of the specimens here examined exhibited this structure, the existence of a penial gland for B. moebii is not confirmed ( Fig. 6 View FIGURE 6 A).
Finally, excluding Baeolidia australis (see B. australis remarks), B. moebii colouration is clearly distinguishable from the remaining species of this genus. In terms of internal anatomy, the radular morphology of Baeolidia moebii is quite particular since teeth are bent and do not have any central notch or denticle. So far, only B. australis and B. lunaris sp. nov. have similar radular teeth. Baeolidia moebii and B. lunaris sp. nov. differ in size ( B. moebii is larger than B. lunaris sp. nov.), and rhinophorial ornamentation ( B. lunaris sp. nov. has elongate papillae instead of minute knobs and colouration). Additionally, the ampulla of B. moebii is larger than the ampulla found in B. lunaris sp. nov. Differences between B. moebii and B. australis are recorded in B. australis remarks.
This species was studied molecularly by Carmona et al. (2013) ( Fig. 1 View FIGURE 1 ).










| MNCN |
Museo Nacional de Ciencias Naturales |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Baeolidia moebii Bergh, 1888
| Carmona, Leila, Pola, Marta, Gosliner, Terrence M. & Cervera, Juan Lucas 2014 |
Baeolidia major amakusana
| Baba 1937: 335 |
Baeolidia moebii
| Eliot 1903: 252 |
| Bergh 1888: 778 |