
Oswaldella vervoorti Peña Cantero & García Carrascosa, 1998
|
publication ID |
https://doi.org/10.11646/zootaxa.4966.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:F4CABF6C-1993-4112-A03E-4DBF708BB025 |
|
DOI |
https://doi.org/10.5281/zenodo.4784055 |
|
persistent identifier |
https://treatment.plazi.org/id/ED10C428-FFF9-2306-FF50-0DE7FB21FD12 |
|
treatment provided by |
Plazi |
|
scientific name |
Oswaldella vervoorti Peña Cantero & García Carrascosa, 1998 |
| status |
|
Oswaldella vervoorti Peña Cantero & García Carrascosa, 1998 View in CoL
( Figs 3–4 View FIGURE 3 View FIGURE 4 )
Oswaldella sp. Peña Cantero, 2009: 1749, fig. 3c.
Material examined. TAN 0602/430, one stem 90 mm high, with gonotheca ( NIWA 144249), one stem 50 mm high, with gonothecae ( NIWA 144253) and one stem 50 mm high ( NIWA 144255).
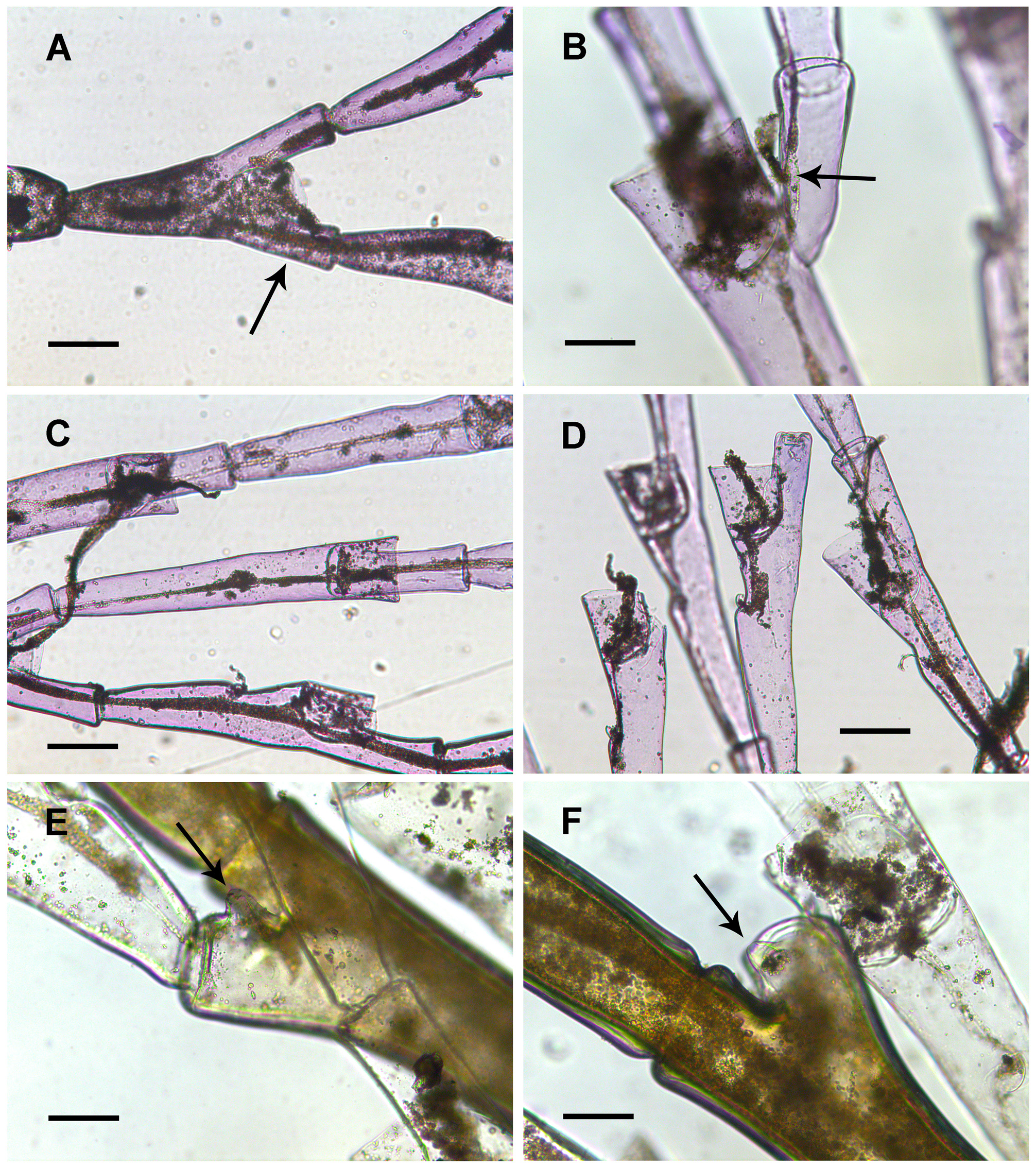
Description. Stems up to 90 mm high, monosiphonic, divided into internodes (one or two apophyses per internode). Cauline apophyses alternately arranged in one plane, with two axillary nematophores and one mamelon ( Fig. 3F View FIGURE 3 ). Cauline apophyses giving rise to hydrocladia with a distinct separation between them. Up to second order hydrocladia present.
Hydrocladia divided into internodes. Unforked hydrocladial internodes ( Fig. 3A–B, G View FIGURE 3 ) with one hydrotheca on distal half of internode and two nematophores: a mesial superior nematophore, placed behind free part of adcauline hydrothecal wall ( Fig. 3E View FIGURE 3 ), and a mesial inferior nematophore, situated on a slightly raised part of the internode and provided with a reduced scale-shaped nematotheca ( Fig. 3C–D, H View FIGURE 3 ). Forked hydrocladial internodes with hydrotheca between two prongs, each of them with a nematophore, and one mesial inferior nematophore provided with a scaleshaped nematotheca. Apophyses supporting lower-order hydrocladia usually smaller than distal part of internodes.
Hydrotheca slightly higher than wide. Adnate to internode for about three-fourths of its adcauline length. Hydrothecal basal part distinctly oblique; abcauline side lower. Abcauline hydrothecal wall longer than adcauline side, aperture slightly directed adcaudally. Distal third of abcauline wall distinctly directed outwards ( Fig. 3C–D, H View FIGURE 3 ).
Gonotheca elongated, club-shaped ( Fig. 3I View FIGURE 3 ).
Measurements (in µm). Hydrotheca: length of abcauline wall from 200 µm at 1 st internode to 300 µm at 7 th, free part of adcauline length 50, diameter at aperture 230–250. Gonothecae: length 2000, maximum diameter 480–700. Nematocysts: larger microbasic mastigophores, 11–12.5 x 3–3.5.
Remarks. One of the 50-mm-high stems (NIWA 144253) is provided with a secondary stem originating from a forked hydrocladial internode at the usual location of the gonothecal insertion. This second-order stem has a typical structure.
Most cauline apophyses of the 90-mm-high stem are deprived of mamelons; a little prominent mamelon was observed only twice. In one of the 50-mm-long stems (NIWA 144255) only some apophyses, particularly at the basal part of the stem, also present mamelon.
In the material studied most hydrocladia are either unforked or just bifurcated (the first internode of primary hydrocladia giving rise to a single secondary hydrocladium). In one of the 50-mm-long stems (NIWA 144255) only seven of its last eight hydrocladia are bifurcated with apophyses quite similar to the distal part of the internode and the hydrotheca approximately in the middle of the bifurcation. There are, however, signs of other branching below. In one case, one first-order hydrocladium has the basal part of four secondary ones, one on each of its four first internodes, resting on distinctly smaller apophyses situated laterally under the hydrotheca. On a few occasions there are secondary hydrocladia that do not originate from the first internodes of the primary hydrocladium, but from others (i.e. there are unforked hydrothecate internodes before the forked ones). This material (NIWA 144255) has hydrothecae ( Fig. 3G–H View FIGURE 3 ) less elongated (abcauline length up to 250 µm) and wider (diameter at aperture up to 280 µm) than those from the remaining material ( Fig. 3A–D View FIGURE 3 ). It also has some nematothecae clearly more developed, with a distinct adcauline wall ( Fig. 3H View FIGURE 3 ). This material, however, is the closest to O. vervoorti by the branching, with unforked hydrothecate internodes between forked ones. According to Peña Cantero & García Carrascosa (1998), although lower-order hydrocladia typically arise from the first internodes of the primary hydrocladia, in many cases unforked hydrothecate internodes were present preceding forked ones. On the other hand, looking at their figures (cf. fig. 1a–c in Peña Cantero & García Carrascosa 1998) it is evident that the apophyses supporting lower-order hydrocladia are smaller than the distal part of the internode. Clearly, it is not a simple bifurcation of the internode. Instead, it is like a normal hydrothecate internode laterally giving rise to the lower-order hydrocladium.
In order to confirm this point, I have examined the holotype of O. vervoorti . The first forked hydrocladial internode is the most misleading as sometimes the apophysis supporting the second-order hydrocladium is similar to the distal part of the internode ( Fig. 4A View FIGURE 4 ); in addition, both the apophysis and the distal part of internode are provided with a nematophore placed at approximately the same level. In all the remaining forked hydrocladial internodes, the prongs are markedly dissimilar ( Fig. 4B View FIGURE 4 ), one is clearly the distal part of the internode, the other is undoubtedly the apophysis. Both are also provided with nematophores, but that of the apophysis is placed at a lower level ( Fig. 4B View FIGURE 4 ). The first internodes originating from the apophyses are relatively very long (up to 1300 µm) ( Fig. 4C View FIGURE 4 ).
The primary hydrocladium might give rise to several second-order hydrocladia (up to six according to Peña Cantero & García Carrascosa 1998). Most secondary hydrocladia originate sequentially from the first (up to the fourth) internodes of the primary one, but it is not unusual to observe extra secondary hydrocladia originating after some unforked internodes (this is clearly shown in figure 1a in Peña Cantero & García Carrascosa 1998).
The first secondary hydrocladium usually gives rise to a third-order hydrocladium at its first internode, but it can also form other tertiary hydrocladia (up to three according to Peña Cantero & García Carrascosa 1998), sometimes also beyond unforked internodes (see also figure 1a in Peña Cantero & García Carrascosa 1998). According to these authors, the first third-order hydrocladium sometimes even forms a fourth-order one.
Mamelons are more prominent in the holotype than in the present material (compare Fig. 3F View FIGURE 3 with Fig. 4E View FIGURE 4 ).
Although suspicious on geographic grounds, there is no evidence to consider the material studied here different from O. vervoorti . This would not be the first Antarctic benthic hydroid previously considered endemic to West Antarctica and later found in East Antarctica.
Peña Cantero (2009) reported, as Oswaldella sp. , material likely conspecific with the one studied here, which is considered to belong to O. vervoorti . The young stems, up to 20 mm high, are divided into internodes with one or two apophyses each, the hydrocladia are unforked and the cauline apophyses are provided with two axillary nematophores and the most distal ones have mamelon. The size of the hydrotheca also agrees.
Ecology and distribution. Present material was collected at depths between 70 and 120 m off Buckle Island; gonothecae in March. The material described as Oswaldella sp. by Peña Cantero (2009) came from a depth of 70–85 m off Buckle Island. Not only is O. vervoorti reported for the first time from the area of study, but also for East Antarctica, which indicate that its geographical distribution may be considered as circum-Antarctic.

| NIWA |
National Institute of Water and Atmospheric Research |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Oswaldella vervoorti Peña Cantero & García Carrascosa, 1998
| Peña Cantero, Álvaro L. 2021 |
Oswaldella sp.
| Pena Cantero, A. L. 2009: 1749 |