
Cryptotropidae Yamaguti, 1958
|
publication ID |
https://doi.org/10.1093/zoolinnean/zlad037 |
|
publication LSID |
lsid:zoobank.org:pub:1D4DD019-F87D-4577-89D3-5C4064085B81C |
|
DOI |
https://doi.org/10.5281/zenodo.10267824 |
|
persistent identifier |
https://treatment.plazi.org/id/EC368786-FFFC-CC59-FC49-FD6CFB72FC6C |
|
treatment provided by |
Plazi |
|
scientific name |
Cryptotropidae Yamaguti, 1958 |
| status |
|
Family Cryptotropidae Yamaguti, 1958 View in CoL
(syn. Renschetrematidae Yamaguti, 1971 )
Diagnosis: Body very small, of varying shape, from ovoid/ rounded and thick to pyriform or linguiform, slender and flattened dorsoventrally. Tegument usually covered almost entirely with small spines, which diminish somewhat in size towards posterior end of body. Suckers well separated; ventral sucker in middle third of body. Suckers can be equal in size or either of them can be slightly larger. Oral sucker subterminal; ventral sucker in middle third of body. Prepharynx absent or very short; pharynx rounded; oesophagus of varying length, from short to long. Caecal bifurcation at various levels between suckers, usually close to ventral sucker; caeca short, terminating approximately at level of oral sucker or posterior to it; in laưer case, not extending past anterior margins of testes.
Two testes, in posterior half of body, post-ovarian, usually very large, opposite or slightly diagonal, longitudinally elongated, with smooth outline or lobed. Copulatory pouch contains winding seminal vesicle, pars prostatica and ejaculatory duct. Copulatory pouch either entirely anterior to ventral sucker or, more rarely, proximal end of copulatory pouch overlapped by ventral sucker or extends past ventral sucker. Distal end of copulatory pouch comma-shaped, curved towards dorsal surface of body. Genital atrium submedian to submarginal, sinistral, usually extracaecal, rarely caecal, between mid-length of oral sucker and ventral sucker. Genital atrium aperture can be surrounded by digitiform spines. Fibrous male accessory pouch (stylet pouch), containing protractible stylet-like rod, can be present in region of ventral sucker, with its distal end opening into genital atrium.
Ovary pretesticular, with smooth outline or somewhat lobed, oval or irregular in shape, usually posterodextral or rarely dextral to ventral sucker. Seminal receptacle usually immediately posterior to ovary. Laurer’s canal and Mehlis’ gland not observed. Vitellarium extensive, in the form of numerous irregularly shaped follicles forming fields of follicles from level of the pharynx to posterior end of body. Characteristically, vitelline fields usually encircle space containing gonads and uterus; in some cases, vitellarium is broken into pretesticular and post-testicular fields or terminates at level of anterior margins of testes. Uterus usually filling the space between ventral sucker and testes and intertesticular space, rarely extending posteriorly somewhat past testes or anteriorly to level of oral sucker. Eggs numerous or relatively few. Eggs can have multiple filaments on opposite poles. In most species, metraterm weakly defined. Excretory pore terminal; excretory vesicle Y-shaped.
In intestines of amphibians, reptiles, birds and mammals; Asia, Africa, North and South America. Life cycle known for one species ( Cephalouterina dicamptodoni ) and involves hydrobiid snails as first intermediate hosts and stoneflies as second intermediate host.
Type genus: Cryptotropa Ozaki, 1926 View in CoL .
A key for identification of cryptotropid genera is provided below.
Key to the genera of the Cryptotropidae Yamaguti, 1958 View in CoL
1a. Fibrous male accessory pouch (stylet pouch), containing protractible stylet-like rod, is present ............................................... 2
1b. Fibrous male accessory pouch (stylet pouch), containing protractible stylet-like rod, is absent ................................................. 4
2a. Genital atrium aperture surrounded by digitiform spines ...................................................................... Armadoatrium View in CoL gen. nov.
2b. Genital atrium aperture is not surrounded by digitiform spines ........................................................................................................ 3
3a. Body nearly round, ovary anterolateral to ventral sucker, testes far apart, uterus extends anteriorly to the level of anterior margin of ventral sucker on both sides .................................................................................................. Rohdetrema Deblock, 2008 View in CoL
3b. Body linguiform, ovary posterolateral to ventral sucker, testes close to each other, uterus postacetabular .................................. ......................................................................................................................................................................... Renschetrema Rohde, 1964 View in CoL
4a. The copulatory pouch is entirely preacetabular or only its proximal end is slightly overlapped by the ventral sucker ........... 5
4b. The copulatory pouch is dorsal to ventral sucker, its proximal end extends posterior to ventral sucker ....................................... ............................................................................................................................................................................ Paracryptotropa View in CoL gen. nov.
5a. Uterus entirely or almost entirely postacetabular .................................................................................................................................. 6
5b. Uterus almost entirely preacetabular, reaching oral sucker ............................................. Cephalouterina Senger & Macy, 1953 View in CoL
6a. Copulatory pouch oriented transversally ............................................................................... Pseudocryptotropa Yamaguti, 1958 View in CoL
6b. Copulatory pouch oriented longitudinally or diagonally ....................................................................... Cryptotropa Ozaki, 1926 View in CoL
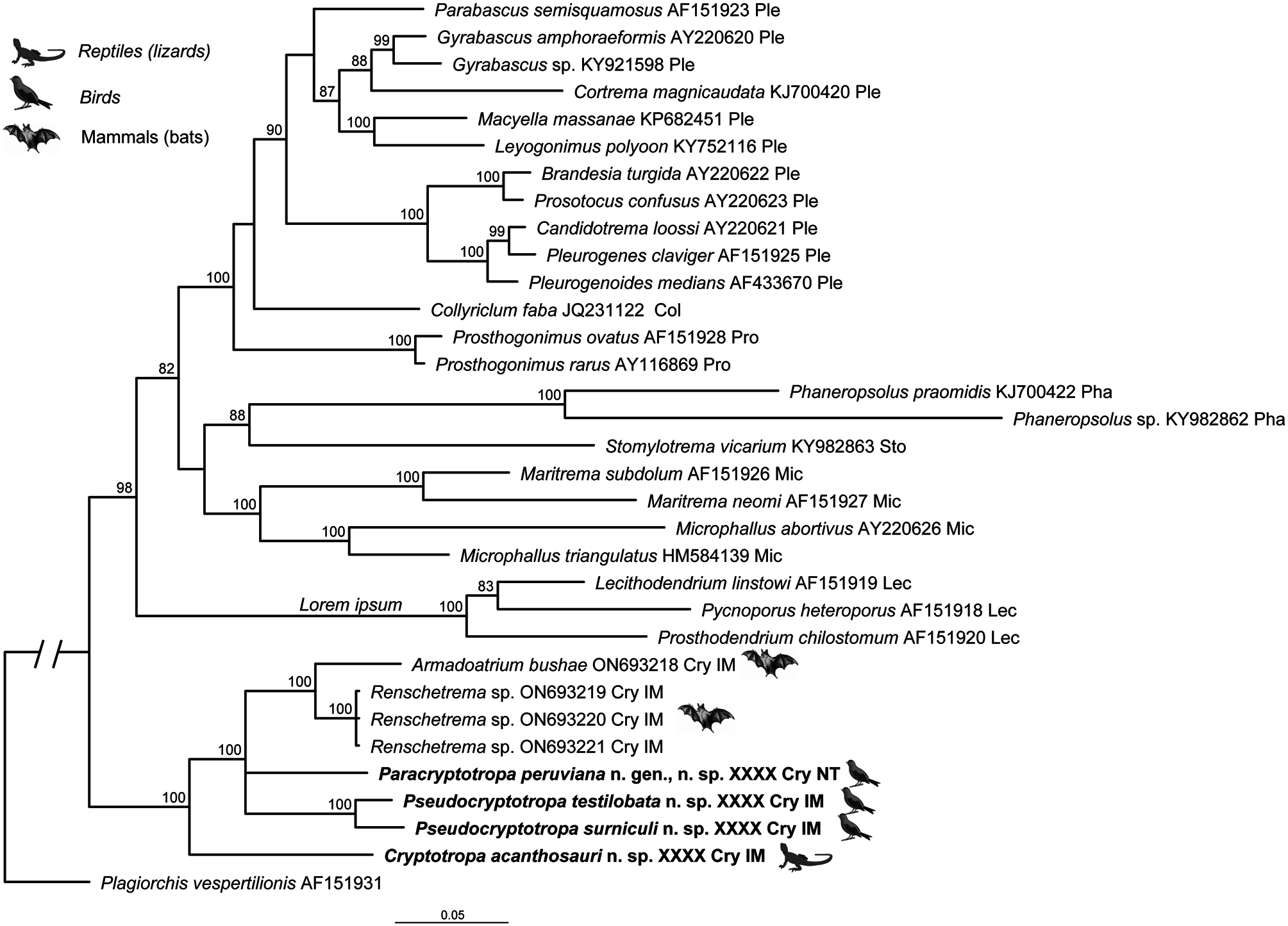
One of the noteworthy results in our molecular phylogeny is the position of the Cryptotropidae View in CoL as the sister taxon to the remainder of the Microphalloidea ( Fig. 1 View Figure 1 ). However, due to the presence of highly unusual and clearly derived characters, such as the dorsal genital pore, in all Cryptotropidae View in CoL , the stylet sac with stylet in Renschetrema View in CoL , Rohdetrema View in CoL and Armadoatrium View in CoL , and the unique egg filaments in Pseudocryptotropa View in CoL , it is difficult to consider the Cryptotropidae View in CoL as the ancestral group or one of the ancestral groups of the Microphalloidea . Considering that a number of microphalloidean taxa have never been included in phylogenetic analyses, there is much to be learned about their interrelationships.
Remarks on morphology
Our examination of freshly collected high-quality specimens has allowed us to address several questions related to the morphology of different members of the Cryptotropidae . One of them was the contradiction between the diagnosis of Cryptotropa in the Keys to the Trematoda (Deblock 2008) and the original description of Cr. kuretanii , regarding the position of the genital pore in this genus. Deblock (2008) wrote that the ‘genital pore opens on anterolateral margin of body’, whereas the original description by Ozaki (1926) said that the ‘genital aperture lies on the dorsal surface, close to the less margin’. This is a critically important difference considering that this feature alone might place a genus into an entirely different family, as has happened in this case. The genital pore in all specimens of our new species of Cryptotropa from lizards in Vietnam is invariably dorsal, although it is positioned close to the less margin of the body.
As mentioned above in the remarks on Renschetrema , our study of high-quality specimens has unequivocally demonstrated the presence of a true copulatory pouch in this genus, as opposed to a free seminal vesicle in parenchyma reported by Deblock (2008). Our examination of the specimens deposited by Klaus Rohde in the collection of the Natural History Museum in London was in agreement with observations of our freshly collected specimens.
Likewise, examination of museum specimens and newly collected material has demonstrated that the statements regarding an independent male and female genital pore in Renschetrema made by previous authors ( Rohde 1964, Matskási 1973, Deblock 2008) have turned out to be erroneous. It is most likely that these authors confused the opening of Laurer’s canal with the female genital pore. Nevertheless, it cannot be excluded that Laurer’s canal in Renschetrema is used for insemination, as was suggested by Khotenovsky (1965) for the Cryptotropidae based on the fact that he observed sperm in the lumen of Laurer’s canal on histological sections but never saw it in the lumen of the uterus.
The polar filaments found in eggs ( Fig. 3B, C View Figure 3 ) of both species of Pseudocryptotropa , described for the first time in the present work, are an extremely unusual feature among microphalloidean taxa. Moreover, the structure of the polar filaments is highly unusual and, to the best of our knowledge, not previously reported in any digenean. Although polar filaments are rare in general, members of several digenean groups (e.g. members of the Notocotylidae Lühe, 1909 and Microscaphidiidae Looss, 1900 ) also have polar filaments. However, in all other digenean groups the filaments, which can be present on one or both poles, are usually whip-like, singular structures (sometimes there is more than one filament on a pole). In Pseudocryptotropa , the filaments appear as multiple fibres of unequal length, with some being glued together with a mucous substance and some not ( Fig. 3B, C View Figure 3 ). It is interesting that despite the high morphological similarity to the two species described herein from the Philippines, the eggs in the type species of the genus, Ps. macrotestis , apparently lack filaments. The redescription of Ps. macrotestis by Khotenovsky (1965) contains a drawing of an individual egg; therefore, we have no reason to believe that the filaments were overlooked. From the functional viewpoint, we hypothesize that the filaments are likely to play a role in facilitating egg swallowing by snail intermediate hosts (e.g. by sticking to vegetation instead of falling to the boưom).
Host associations and geographical distribution
Our new host and geographical records have documented the extremely broad geographical distribution of the Cryptotropidae and a high level of evolutionary ecological/physiological plasticity of these digeneans to their hosts. The Cryptotropidae are now reported from amphibians (in North America), reptiles (in Southeast Asia and Africa), birds (East and Southeast Asia and South America) and mammals (Southeast Asia) ( Fig. 1 View Figure 1 ; Table 1 View Table 1 ). Members of only a few digenean families are found in such a broad array of hosts that includes both cold- and warm-blooded vertebrates. This is remarkable considering that the Cryptotropidae are a small family, with only seven genera and 13 species, including the four new species described herein. It is noteworthy that a member of Cryptotropa , a genus including only parasites of amphibians and reptiles, appears to be basal to taxa from birds and mammals in both the 28S and cox1 trees ( Figs 1 View Figure 1 , 2 View Figure 2 ). The 28S data also suggest that the two genera from bats, Renschetrema and Armadoatrium , are closely related to each other, which is also supported by their morphological similarity. However, drawing definitive conclusions regarding the evolution of host associations or biogeography in this family based on the currently available dataset would be premature.
Despite the relatively low taxonomic diversity of cryptotropids (13 species in seven genera) currently known, they occur on four continents and from an unusually broad range of definitive hosts (amphibians, reptiles, birds and small mammals). The two new species of Pseudocryptotropa described herein are the first members of the Cryptotropidae from avian hosts reported in the Philippines. Paracryptotropa peruviana is the first representative of the Cryptotropidae in South America and only the second species of the family known from the Western Hemisphere. The other species is Ce. dicamptodoni , described from caudatan amphibians in the Pacific Northwest of the USA. Cryptotropa acanthosauri is the first member of the genus from a reptilian host in Asia.
Thus, only a single cryptotropid species is currently known from Africa, North America and South America. This paưern is more likely among parasites of ancient host groups characterized by very limited diversity and/or limited distribution or dispersion ability (e.g. crocodilians). However, cryptotropids are found in highly diverse and mostly evolutionarily derived groups of vertebrate hosts, such as anuran amphibians, lizards, passeriform, cuculiform and apodiform birds, bats and primates. Given the fact that only three cryptotropid species are currently known outside Asia, combined with the extremely diverse fauna of potential definitive hosts, especially in tropical and subtropical regions of the world, we posit that the real diversity of the Cryptotropidae is substantially greater than currently known, and there is liưle doubt that many new taxa belonging to this unique group of digeneans await discovery.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Digenea |
|
Order |
|
|
Family |
Cryptotropidae Yamaguti, 1958
| Tkach, Vasyl V., Chermak, Taylor P., Patitucci, Kaylyn K., Greiman, Stephen E., Binh, Tran Thi & Olson, Peter D. 2023 |
Renschetrematidae
| Yamaguti 1971 |