
Heliotrygon
|
publication ID |
https://doi.org/ 10.5281/zenodo.276900 |
|
DOI |
https://doi.org/10.5281/zenodo.5617656 |
|
persistent identifier |
https://treatment.plazi.org/id/EB76AC73-E94D-0041-7781-F8FAFB5BE5D1 |
|
treatment provided by |
Plazi |
|
scientific name |
Heliotrygon |
| status |
|
Morphology of Heliotrygon View in CoL View at ENA
The description of dermal denticles that follows is based on H. rosai (MZUSP 108295), but the ventral lateral-line canals are based on H. gomesi (MZUSP 108203). Clasper skeleton is described based on adult claspers of H. rosai (MZUSP 108295), as adult claspers of H. gomesi were not available for dissection, but clasper external morphology (included above in the account on external morphology of H. rosai ) was based on the holotype (MZUSP 104996). Description of cranial muscles is also based on H. rosai (MZUSP 108295).
Dermal denticles. ( Figures 9 View FIGURE 9 A, 18, 23A, 23C). Denticles small (much smaller than eye-diameter), evenly distributed, and present on dorsal disc region mostly over mid- and posterior disc and base of tail. Denticles very minute and low. Larger spines and thorns entirely absent. On tail region, denticles more intensely grouped together at tail base, with scattered denticles occurring on lateral tail aspects, and sporadically far distally on tail. Microscopically, dermal denticles on dorsal disc region of a single morphotype, with an oval-shaped crown and averaging around 0.5 mm in length. Posteriorly on the crown, two parallel crown keels originate from a coronal dichotomy. The opposite side of the crown is round to oval. Denticle basal plate oval, interrupted in the coronal keel area. A small basal keel occurs in variable numbers, being sometimes absent. In lateral view, denticle base and basal keels visible. Denticles on dorsal and lateral tail regions with slightly different morphology from dorsal middisc ( Figure 9 View FIGURE 9 A), star-shaped in dorsal view, and with radiating keels converging on denticle apex.
Ventral lateral-line canals. ( Figure 19 View FIGURE 19 ). Ventral lateral-line canals present a complex, honeycomb-like pattern on anterolateral disc region. Suborbital component of the infraorbital canal highly reticulated and web-like, delimiting a network of small loops of varying sizes. Suborbital component covering most of anterolateral disc region, from level of fourth gill opening to outer anterolateral disc margin. Loops of suborbital greater and more elongated posteriorly, reducing in size outwardly toward disc margins, becoming very small close to anterolateral disc margins near hyomandibular canal. Hyomandibular canal subdivided into jugular and subpleural components. Hyomandibular canal bordering anterolateral disc margins, running more or less parallel to disc margin. Subpleural component of the hyomandibular canal deflects inward toward disc center more or less at level of third gill opening, where it delimits irregular circular to oval loops. Subpleural extends posteriorly to posterior one-fourth of disc. Anterolaterally the hyomandibular canal gives off numerous small, perpendicular branches toward disc outer margin (the anterior subpleural tubules). Supraorbital canal extends from nasal curtain region posteriorly before deflecting to form slender, elongated anterior loop. Posteriorly, supraorbital canal reaches slightly caudal to level of mouth. Prenasal canal extends anteriorly in a more or less parallel fashion from nasal curtain to anterior disc. The jugular component of the hyomandibular canal reaches posteriorly, arching around branchial basket from junction with suborbital and supraorbital canal, to reach subpleural component at posterior three-fourths of disc.
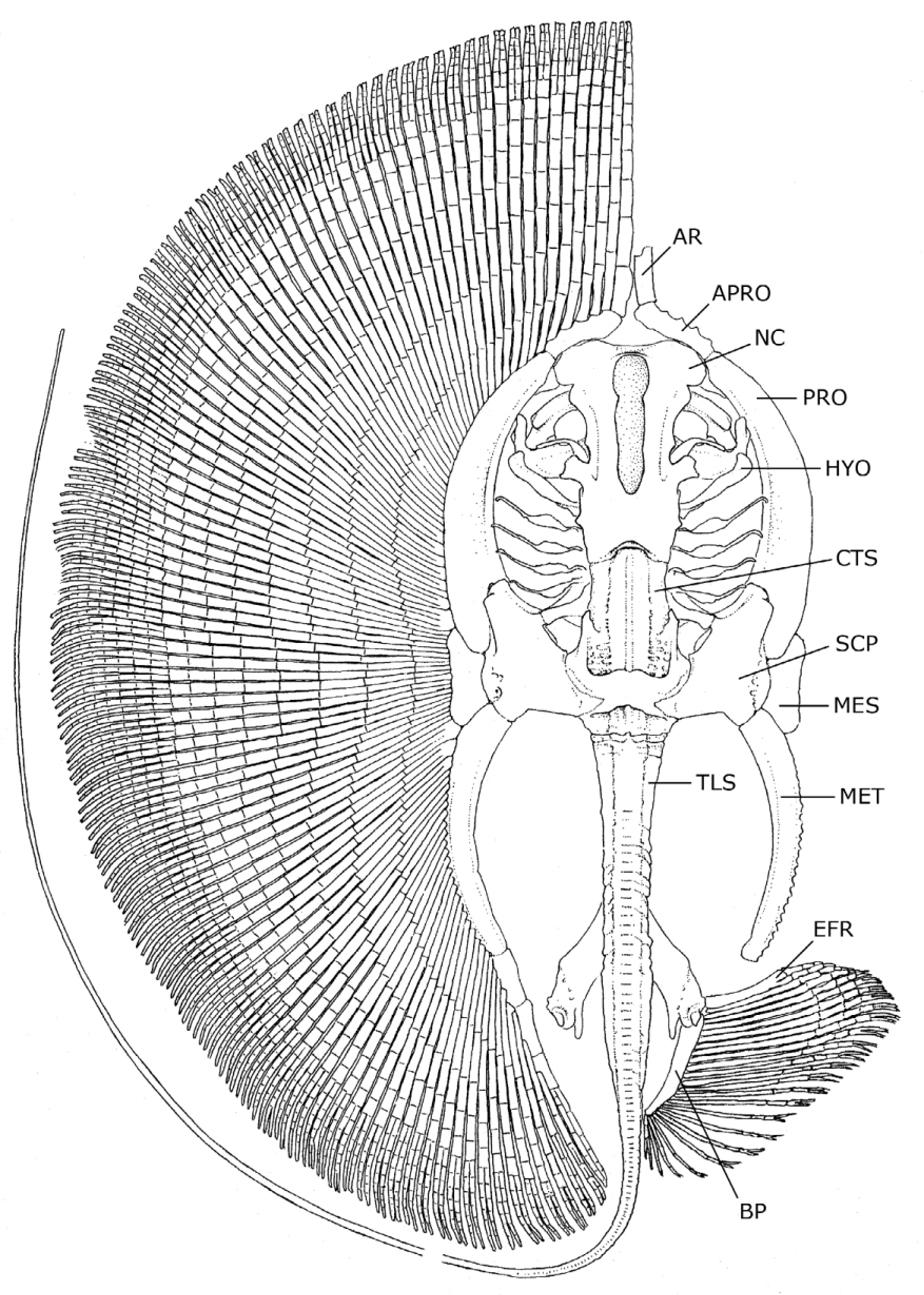
Skeletal anatomy. ( Figures 20–4 View FIGURE 20 ). In general, the skeleton of the new genus (based on examinations of both species) is very similar to that of Paratrygon . The description below roughly applies to both H. gomesi and H. rosai , and was based on specimens of both species.
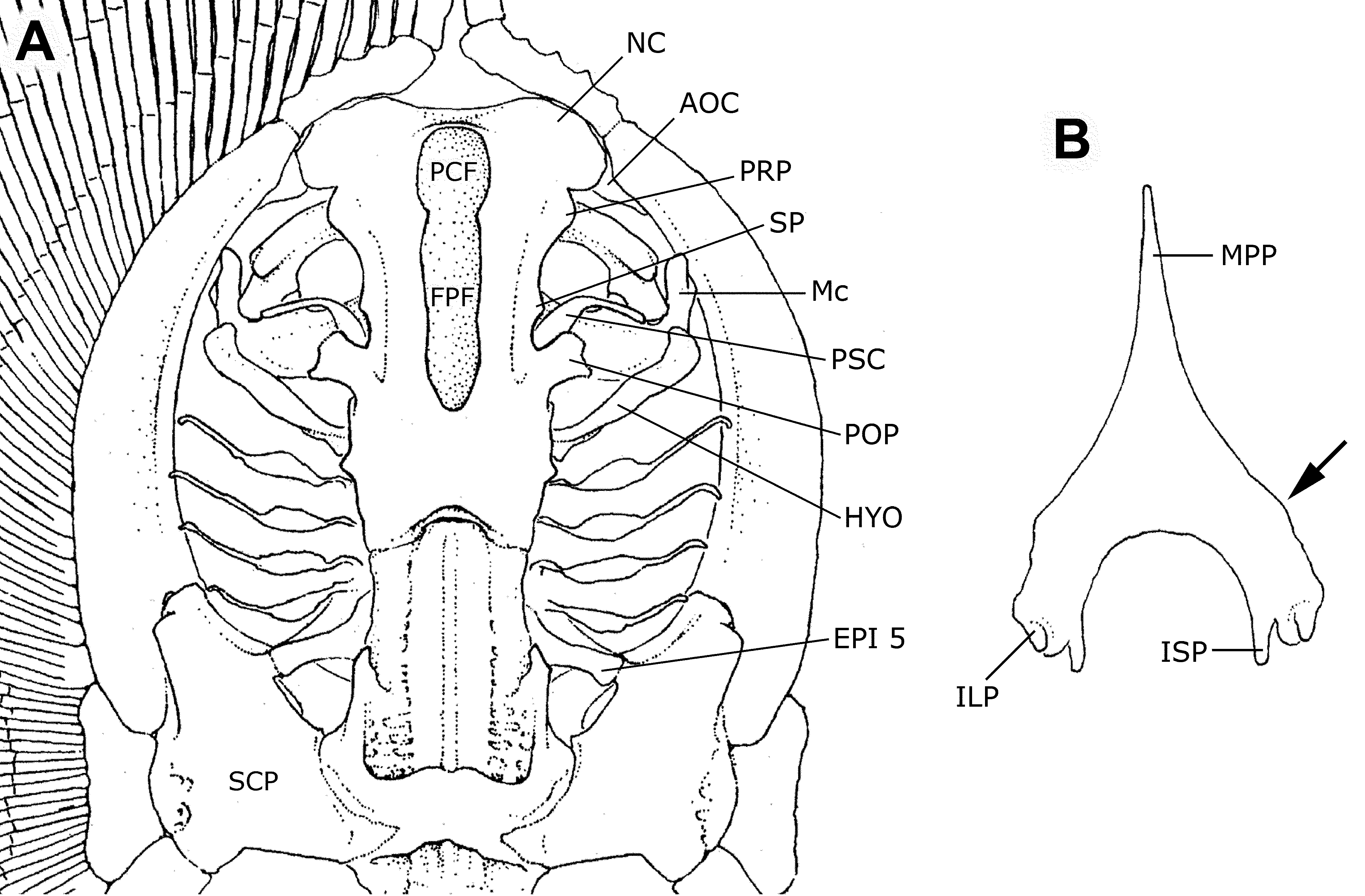
Neurocranium. The neurocranium is relatively elongate, being about as long as the metapterygium, and twice as long as neurocranial greatest width. Neurocranial greatest width at nasal capsule region. Nasal capsules broadly rounded anteriorly and relatively short anteroposteriorly. Postorbital processes wider than preorbital processes. Precerebral and frontoparietal fontenellae elongate, about three-fourths of neurocranial length, and composed of a single structure in adults (no epiphysial bridge present). Precerebral fontanelle extends anteriorly to almost neurocranial anterior margin. Post-fontanellae neurocranial length relatively great, about one-third of neurocranial length. Nasal capsules separated by a relatively wide internasal septum, about equal to nasal aperture length. Anterior margin of nasal capsules slightly irregular, with small protuberances medially and at outer corners, articulating with anterior segment of propterygium. No sign of rostral appendix or remnant of rostral process in any specimen. Between pre- and postorbital process, much closer to the postorbital process, is the small, triangular supraorbital process. Preorbital processes small and blunt. Spiracular cartilage (providing support for the anterior spiracular wall) slender and elongate, curved at more or less its midlength. Antorbital cartilage triangular, articulating with posterolateral corner of nasal capsule, and closely associated with propterygium.
Mandibular, hyoid and branchial arches. In general, the mandibular arch resembles that of Paratrygon , but slightly less angled and more straight across. Palatoquadrates very slender and relatively straight at anterior margins. Meckel’s cartilages slightly oriented anteriorly at mid-line, but generally also relatively straight. Meckel’s cartilages more robust at symphysis, and with well-developed, slender lateral processes. Hyomandibulae more or less straight and relatively short, anteriorly slightly curved toward midline, with small anterior concavity for articulation with Meckel’s cartilages. Angular cartilages absent. Basibranchial copula elongate, reaching anteriorly to beyond posterior margin of frontoparietal fontanelle in dorsoventral view, and extending to close to basihyal cartilage. Anterior segment of basibranchial very triangular, acute, and more slender than in other potamotrygonid genera. First hypobranchial elements anteriorly oriented toward midline, extending anteriorly to basibranchial copula extremity. Pseudohyoid arch more slender than succeeding branchial arches. Branchial gill rays reaching propterygium.
Axial elements. Cervicothoracic synarcual cartilage very robust and elongate, its width anteriorly about equal to posterior neurocranial width, and its greatest width just under greatest width of neurocranium. Lateral stays not projecting laterally to a great extent. Thoracolumbar synarcual strongly calcified laterally and relatively wide, extending from scapulocoracoid to close to puboischiadic bar (posterior to anterior extremity of median prepelvic process). Individual vertebral elements occurring posteriorly beyond level of caudal sting, becoming an uncalcified notochordal extension (cartilaginous rod) well posterior to caudal sting extremity. Transition from mono- to diplospondyly occurs usually slightly caudal to posterior margin of pelvic girdle (usually at between third and fifth centrum posterior to pelvic girdle). Distinct neural elements present even on vertebrae of caudal whip.
Appendicular skeleton. Propterygium relatively stout, widest posteriorly, and anteriorly curved toward midline. Anterior segment of propterygium about one-fourth to one-fifth length of propterygium, closely abutting nasal capsule, and almost contacting its antimere. Mesopterygium also relatively elongate, slightly concave externally, and more slender anteriorly. Anterior and posterior extensions of mesopterygium rounded. Metapterygium highly arched, more slender than propterygium, articulating posteriorly with three smaller metapterygial segments. Disc with closely articulating pectoral fin radial elements. Radial elements sometimes fused at base, especially those articulating with posterior portion of propterygium and anterior segment of metapterygium. Pectoral radials relatively slender where they articulate with pectoral basals, widening toward middisc, and becoming again more slen- der distally where they bifurcate. Pectoral radials subdivided into 17 segments (18 in some specimens) from propterygium to outer disc margin; complete subdivision of radials occurs at ninth segment at level of greatest disc width, but at eighth segment closer to anterior disc. Radial subdivision occurs by splitting and strengthening internal calcification in segment medial to segment that is fully subdivided. Radial segments more stout at anteriormost disc on snout region, and becoming progressively shorter toward outer disc margins. Outermost radials with distal subdivided segments closely contacting each other.
Pelvic girdle very triangular in profile, relatively very stout, with median prepelvic process extending to about anterior one-third of metapterygial length (extent of prepelvic process not visible in radiographs) ( Figure 22 View FIGURE 22 B). Lateral prepelvic processes reduced to mere rounded corners. Pelvic girdle resembling an inverted “V”, with short iliac processes not extending caudally beyond ischial processes. Iliac processes with articular surface for basipterygium and enlarged first pelvic radial element. Ischial processes not very elongate, slender distally. Basipterygium robust, about as wide as posterior portion of metapterygium. Basipterygium curves inward posteriorly, and oriented obliquely in relation to midline. Pelvic radials subdivided into eight or nine segments, the most proximal much more elongate, extending to about 80% of pelvic fin width; fifth pelvic segment is subdivided.
Clasper skeleton ( Figure 24 View FIGURE 24 ) consists of the following cartilages: two basal segments, a single beta cartilage, axial cartilage, dorsal marginal, ventral marginal, dorsal terminal 2, accessory terminal, and ventral terminal. First basal segment larger and connecting to basipterygium; second basal segment smaller and linked to proximal part of axial cartilage. Beta cartilage originating at first basal segment and distally articulated with dorsal marginal. Axial cartilage straight, depressed anteriorly and distally cylindrical, tapering toward extremity. Ventral marginal long and narrow, with pronounced anterior tip. Two pelvic radials reach anterior tip of ventral marginal (not depicted in illustration, Figure 24 View FIGURE 24 ). Outer edges of dorsal marginal and dorsal terminal 2 forming the clasper groove externally. Inner edge of posterior portion of dorsal marginal forming dorsal pseudosiphon externally. Dorsal terminal 2 elongated, narrow and oval. Accessory terminal cartilage elongated and fusiform, underlying dorsal terminal 2, and forming ventral pseudosiphon externally from its outer edge. Ventral terminal broad, elongate and oval.
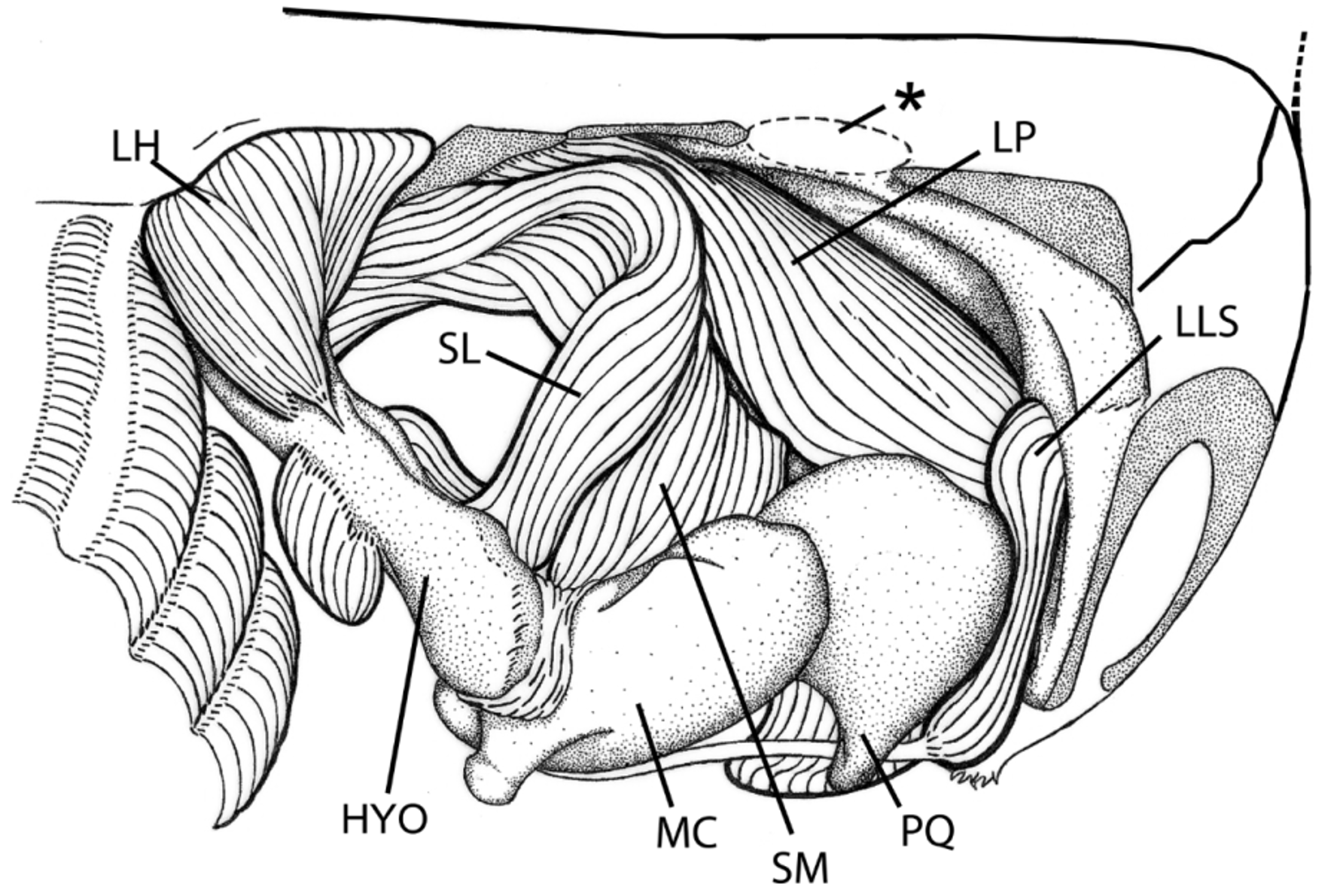
Cranial, hyoid and mandibular muscles. ( Figure 25 View FIGURE 25 ). A full description of the myology of Heliotrygon is being prepared; here we restrict ourselves to features that clarify the affinities of Heliotrygon with other potamotrygonids. Heliotrygon has a considerably robust mandibular musculature. The short distance between the neurocranium and the branchial region has confined the ventral hyoid muscles to a relatively small area, very similar to the condition in Paratrygon . In terms of cranial muscles, Heliotrygon has other features in common with Paratrygon , hypothesized to be derived: (1) the lateral adductor mandibulae 1 has an elevated number of superficial subdivisions (muscle fibers are internally somewhat mixed); (2) medial portion of spiracularis (spiracularis medialis) is inserted onto Meckel’s cartilage, and does not project ventrally as in Plesiotrygon and Potamotrygon . Spiracularis lateralis muscle inserts onto distal portion of the hyomandibula. Levator palatoquarati muscle wraps around palatoquadrate to insert onto its outer, medial portion. The coracohyoideus muscle shows two muscle fibers oriented in different directions, however an evident division into two separate muscles (as in Paratrygon and in some Potamotrygon species) is not visible (in Plesiotrygon , all the fibers have the same orientation).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Chondrichthyes |
|
Class |
|
|
Order |
|
|
Family |