
Centris
|
publication ID |
https://doi.org/ 10.5281/zenodo.187210 |
|
DOI |
https://doi.org/10.5281/zenodo.6213795 |
|
persistent identifier |
https://treatment.plazi.org/id/E92ED043-FFA8-C510-FF0E-FC55B4A8FF2E |
|
treatment provided by |
Plazi |
|
scientific name |
Centris |
| status |
|
Species of Centris View in CoL of the hyptidis group
The species of the hyptidis group present unique characters within Centris , particularly in the morphology of the elaiospathes, which includes the extremely small posterior giant setae (secondary comb), apparently not used for oil collection, judging by their vestigial condition. This group of species presents interesting associations with oil-producing plants, mainly of the genus Angelonia (Plantaginaceae) , which possess trichomatic elaiophores, which, unlike the epithelial elaiophores, require specialized structures for their exploitation (Vogel 1974; Neff & Simpson 1981; Simpson et al. 1990). Due the positioning of the elaiophores in these plants, often arranged within a pair of sacks or cavities, they are accessible only to the first pair of legs of bees, which present the setae of the primary comb of the midlegs reduced or separated and, therefore, no longer used for the oil-collection ( Vogel & Machado 1991). Such behavior, correlated with its morphofunctional complement, seems to be an apomorphy within Centridini ( Vogel & Machado 1991), in which females are basically equipped to oil-collection activity using fore and midlegs ( Neff & Simpson 1981; Vogel & Machado 1991), with both pairs of legs possessing compact and well-developed setal combs.
Given the location of the elaiophores in Plantaginaceae , oil gathering is restricted to a limited number of visitors, which as in the case of the females of the hyptidis group, possess their elaiospathes highly modified not only by the reduction of the secondary posterior setae but also by the presence of long and branched bristles of spatulate apex ( Figures 9 View FIGURES 9 – 12 C and G in Vogel & Machado 1991). These bristles are used for oilcollection not through scraping of the elaiophores as in other species of Centris , but by absorption, using capillarity forces ( Vogel & Machado 1991).
In the case of Centris hyptidis , it is observed a significant reduction in the size of the setae forming the primary anterior comb, which remains almost hidden among the rest of the pilosity of the basitarsi. Fore legs of C. hyptidis are disproportionately long, which allow access to elaiophores of e.g., Angelonia pubescens , which is visited only for oil gathering, being in turn, its unique legitimate visitor ( Vogel & Machado 1991). On the other hand, females of Centris hyptidoides and C. thelyopsis n. sp. possess forelegs with normal length and the primary comb less reduced.
Within Centridini, there is an ancestral relationship with plants with epithelial elaiophores, particularly with species of Malpighiaceae . Independent losses of the ability to collect oils, with a corresponding reduction or loss of the respective structures have been observed in several species-groups and subgenera (for example in C. (Wagenknechtia) moldenkei Toro & Chiappa , some species of C. (Penthemisia) and species-groups in North and South American C. (Paracentris )), along with a variety of modifications of the oil-collecting apparatuses, related with the radiation in the spectrum of oil hosts.
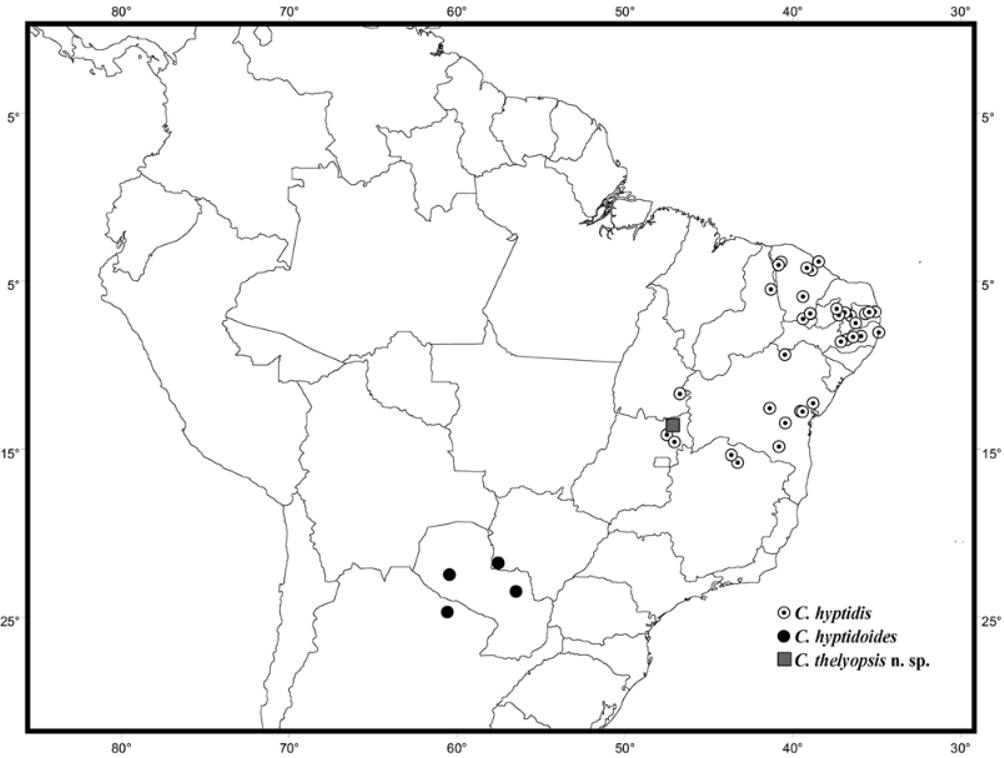
The species of the hyptidis group are associated with the diagonal of open dry areas, with C. hyptidis in the Caatinga (including peripheral areas in the upper Tocantins basin, in northeastern Goiás, Brazil), C. hyptidoides in the Chaco, and C. thelyopsis n. sp. in the dry forests of the upper Tocantins river ( Fig. 13 View FIGURE 13 ). Several cases of bee groups with endemism in both the Chaco and Caatinga are well-known (e.g., Zanella 2002a; Ramos & Melo 2007). None of these groups, however, contain species endemic to the dry forests of the valley of the Paranã river and slopes of the ‘Serra Geral’, in the upper Tocantins basin, in northeastern Goiás. Other bee species considered endemic to the Caatinga domain, e.g., Arhysoceble huberi (Ducke) and Caenonomada unicalcarata (Ducke) , or to smaller areas within it, e.g. Camargoia nordestina Camargo , have also been collected at the type locality of C. thelyopsis n. sp. in the Paranã valley (G. Melo & A. Aguiar, unpublished data). Silva (1997) and Silva and Bates (2002), in studies of bird distribution patterns within the Brazilian Cerrado, have recognized the Paranã valley as an area of endemism, based on the distribution of the two bird species, Knipolegus franciscanus Snethlage (Tyrannidae) and Pyrrhura pfrimeri Miranda-Ribeiro (Psittacidae) . The first bird species is also found in the dry forests of the middle São Francisco valley, in Bahia and northern Minas Gerais ( Silva & Oren 1992), and therefore, has a distribution pattern comparable to that of C. hyptidis , while the psittacid is restricted to northeastern Goiás ( Olmos et al. 1997).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.