
Byrsopteryx Flint, 1981
|
publication ID |
https://doi.org/ 10.5281/zenodo.276427 |
|
DOI |
https://doi.org/10.5281/zenodo.6211681 |
|
persistent identifier |
https://treatment.plazi.org/id/E862879F-FF80-EE29-FF1B-3923FC81FE57 |
|
treatment provided by |
Plazi |
|
scientific name |
Byrsopteryx Flint, 1981 |
| status |
|
Genus Byrsopteryx Flint, 1981 View in CoL
Byrsopteryx Flint 1981: 27 View in CoL [type species: B. mirifica Flint, 1981 View in CoL , original designation]; Harris & Holzenthal 1994 [revision, placement]; Bowles et al. 1999: 45 [placement].
Byrsopteryx carioca sp. nov. ( Figs. 1–21 View FIGURES 1 – 3 View FIGURES 4 – 7 View FIGURES 8 – 9 View FIGURES 10 – 15 View FIGURES 16 – 17 View FIGURES 18 – 27 )
Diagnosis. The male of Byrsopteryx carioca sp. nov. resembles that of B. abrelata in many respects. Males of both species have an elongate sternum VIII, elongate dorsolateral processes on segment IX, each narrowing distally to an acute downturned apex, and the subgenital plate is wide basally and narrow apically in ventral view, with a mesal projection. Males of the new species can be distinguished from those of B. abrelata and other species in the genus by segment IX possessing a pair of ventral, stout, spine-like processes posteriorly, by the dorsolateral processes from segment IX each with a basal acute ventral projection when viewed laterally, and by the apex of the phallus bearing internally a short bifid sclerite.
Females of the new species can be distinguished from those of other Byrsopteryx species by segment VI lacking a ventromesal process, by segment VIII being almost as long as wide, and by the sclerotized, anteriorly rounded vaginal apparatus with a pair of lightly sclerotized lateral arms posteriorly.
Byrsopteryx View in CoL larvae were previously described for B. mirifica View in CoL and B. bipartiterga Botosaneanu, 2000 . Larvae of Byrsopteryx carioca sp. nov. are very similar to those of B. mirifica ( Holzenthal & Harris 1992) View in CoL . However, in B. carioca the abdominal segments of the larvae are not laterally compressed as in B. mirifica View in CoL and the case is slightly depressed dorsoventrally. We cannot distinguish larvae of the new species from B. bipartiterga because few features were provided in the original larval description ( Botosaneanu 2000).
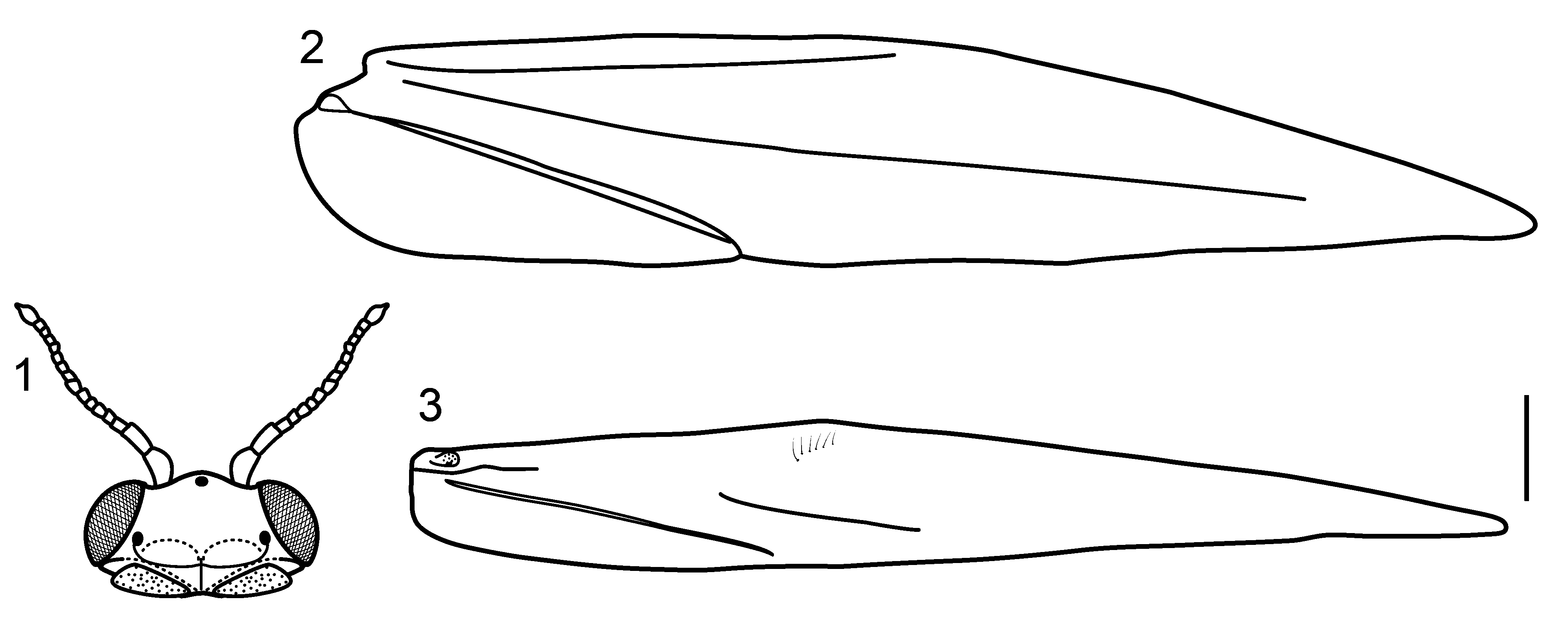
Description. Holotype male. Length 2.4 mm. Head dark brown, eyes reddish brown; antennae simple, short, 16-articulated, scape elongate, with internal margin convex, pedicel elongate, terete, flagellomeres I- XIII short, flagellomere XIV oval with small apical projection ( Fig. 1 View FIGURES 1 – 3 ); frontal region with white setae; 3 ocelli; maxillary palpi 5-articulated; labial palpi 3-articulated. Mesoscutellum with transverse suture. Forewings black with 3 distinct white spots, basal and apical spots small, median spot large, venation reduced, with distinct “line of weakness” ( Fig. 2 View FIGURES 1 – 3 ); hind wings with hooked setae near costal margin, venation reduced ( Fig. 3 View FIGURES 1 – 3 ). Legs dark brown, covered by short black setae; tibial spur formula 0,3,4. Abdomen pale brown; segment VII without ventromesal process.
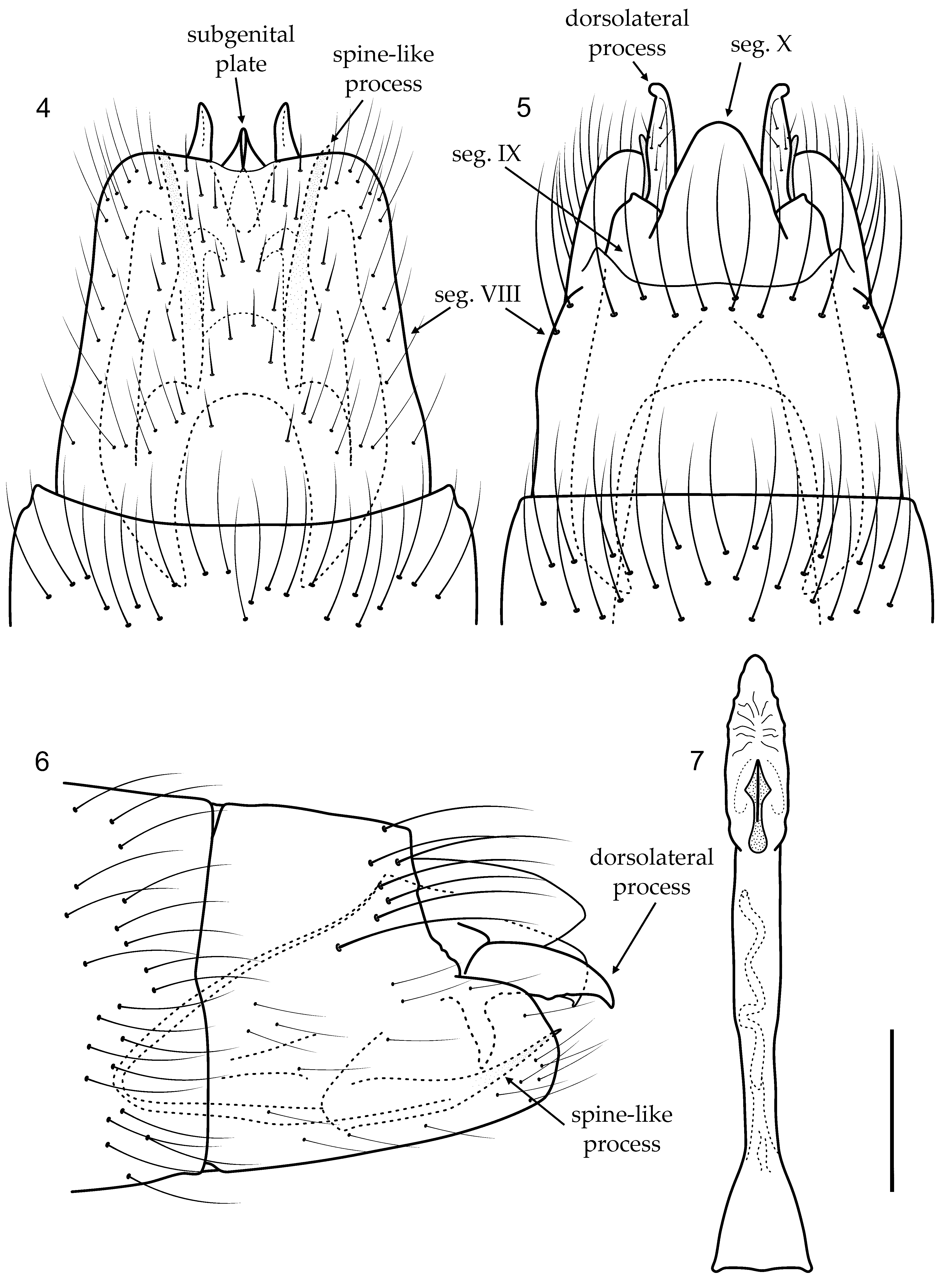
Male genitalia. Segment VIII with sternum longer than tergum; in ventral view, bearing short scattered setae, posterior margin rounded, with mesal area slightly sclerotized forming short rounded incision ( Fig. 4 View FIGURES 4 – 7 ); in dorsal view, with row of long setae near posterior margin ( Fig. 5 View FIGURES 4 – 7 ). Segment IX recessed within VIII, narrowing anteriorly in lateral view, with deep mesal incisions anteriorly in ventral and dorsal views; with pair of stout spine-like processes posteriorly ( Figs. 4, 5, 6 View FIGURES 4 – 7 ); dorsolateral process narrowing to acute, downturned apex, with basal acute ventral projection, in lateral view ( Fig. 6 View FIGURES 4 – 7 ); in dorsal view, apex of dorsolateral process curved laterad ( Fig. 5 View FIGURES 4 – 7 ). Inferior appendages not apparent. Subgenital plate, in ventral view, wide basally, narrowing to apex, with mesal projection ( Fig. 4 View FIGURES 4 – 7 ); in lateral view, truncate, with small apical projection ( Fig. 6 View FIGURES 4 – 7 ). Phallus simple, basal area wide, mesal area slightly constricted, but almost cylindrical, ejaculatory duct sinuous, apical area membranous, internally with short securiform sclerite, divided apically ( Fig. 7 View FIGURES 4 – 7 ).
Va r ia ti o n. Length 2.4–2.6 mm (n=3). Other features of the paratypes similar to the holotype.
Female. Length 3.0– 3.1 mm (n=2). Coloration and features of head and thorax as in male. Abdominal segment VI without ventromesal process.
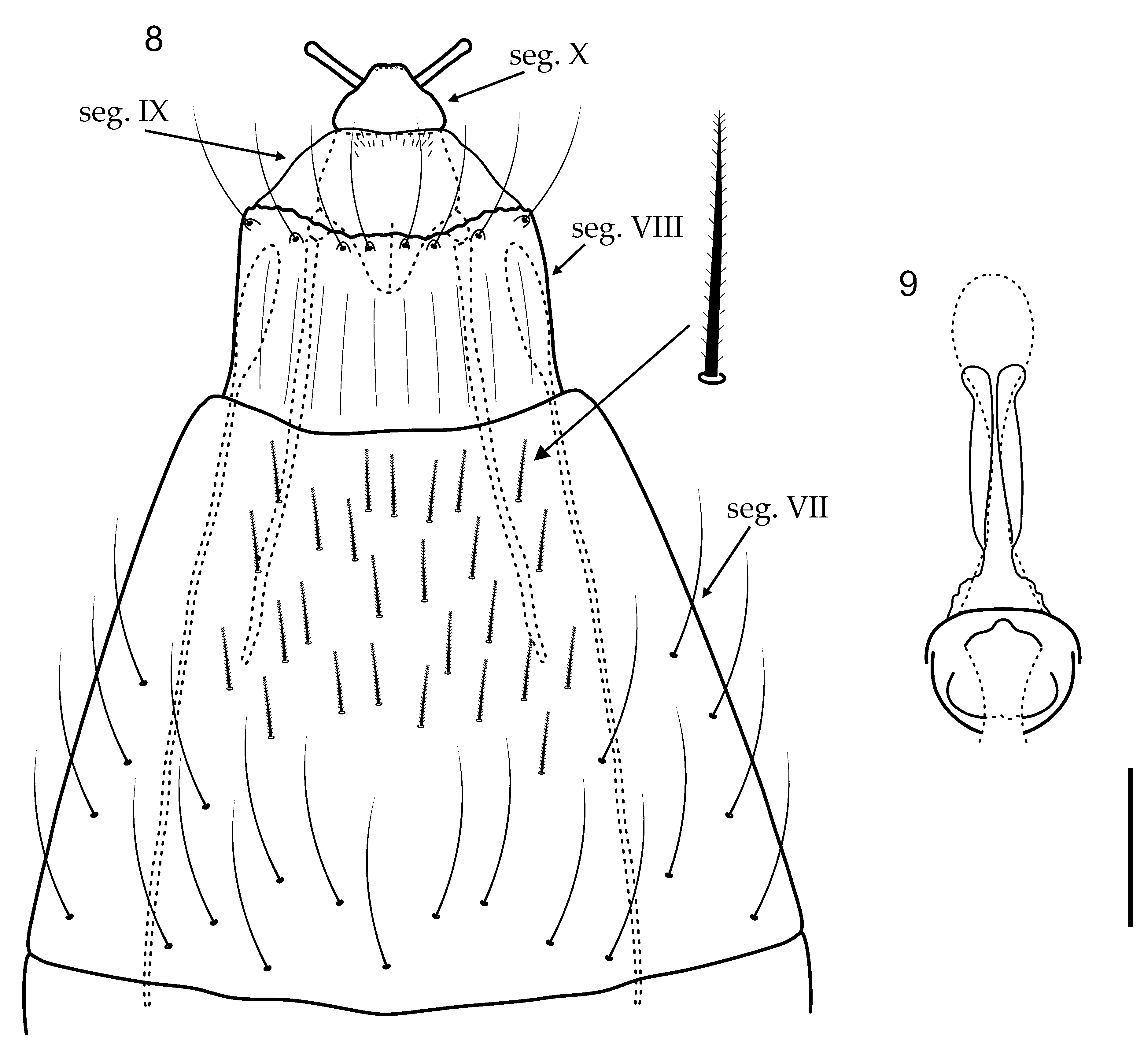
Female genitalia. Segment VII elongate, narrowing posteriorly, ventrally covered by short plumose setae ( Fig. 8 View FIGURES 8 – 9 ). Segment VIII as long as wide, posterior margin slightly emarginate with ring of stout setae; internally with a pair of elongate lateral apodemes extending to posterior of segment VI ( Fig. 8 View FIGURES 8 – 9 ). Segment IX short, internally with a pair of elongate lateral apodemes extending to middle of segment VII ( Fig. 8 View FIGURES 8 – 9 ). Segment X short, wide basally narrowing apically, bearing a pair of papillae ( Fig. 8 View FIGURES 8 – 9 ). Vaginal apparatus sclerotized anteriorly, with anterior margin rounded in ventral view; posteriorly with 2 lightly sclerotized lateral arms ( Fig. 9 View FIGURES 8 – 9 ).
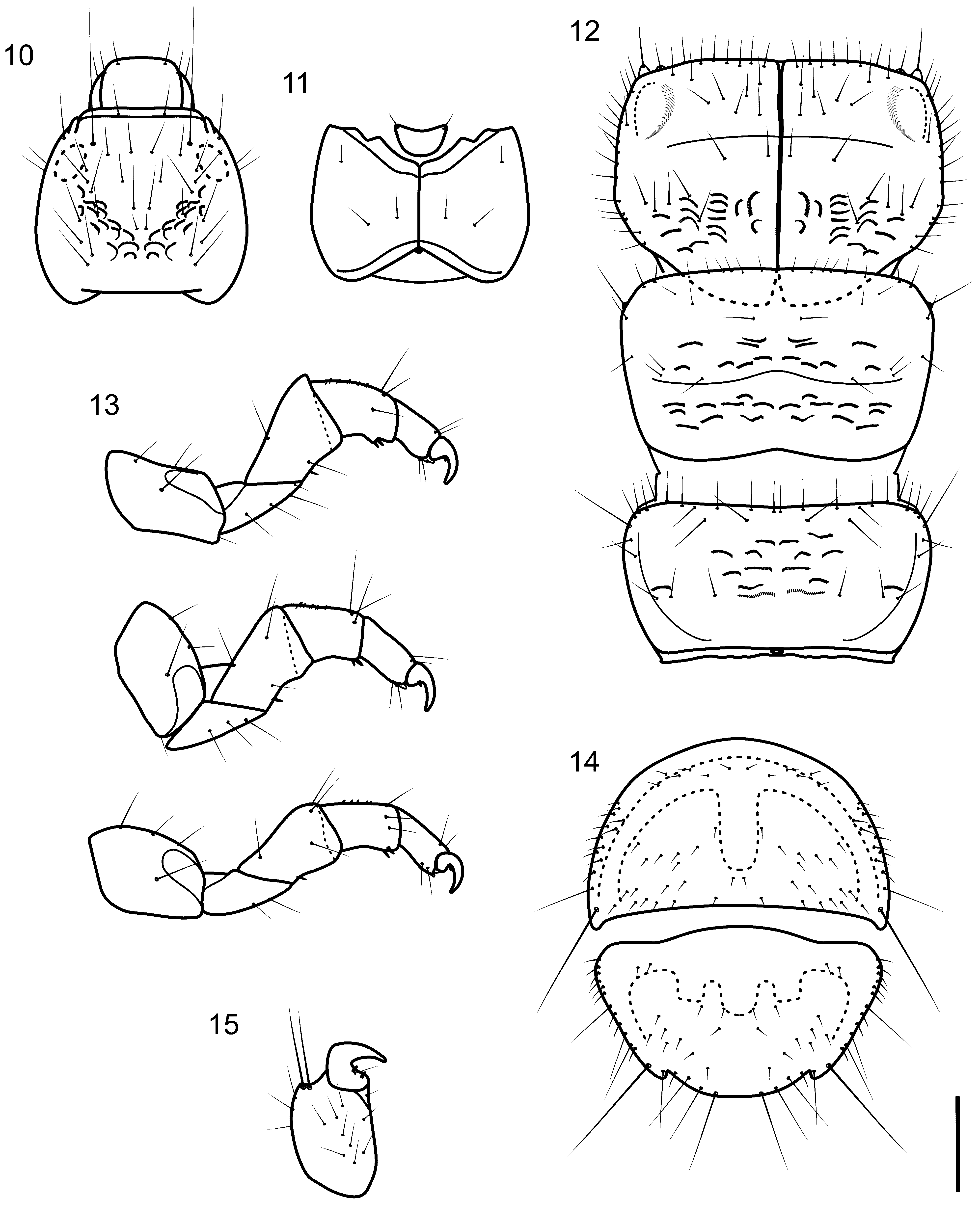
Larva (final instar). Length 2.4–3.8 mm (n=14). Head dark brown, unpigmented around eyes; quadrangular ( Fig. 10, 11 View FIGURES 10 – 15 ); frontoclypeal and coronal sutures indistinct; dorsum with reticulate pattern formed by microscopic setae; setae 9 very long ( Fig. 10 View FIGURES 10 – 15 ); antennae 1-articulated, apparently without apical seta; mandibles without distinct teeth. Thoracic nota strongly sclerotized, dark brown, with setae, particularly on margins, and microscopic setae present; pro- and mesonota with transverse mesal sclerotized ridge, inconspicuous on metanotum; pronotum with middorsal ecdysial line, pair of anterolateral depressed areas ( Fig. 12 View FIGURES 10 – 15 ); meso- and metanota slightly more broad than long, without middorsal ecdysial lines ( Fig. 12 View FIGURES 10 – 15 ); meso- and metathoracic pleura large, sclerotized; pair of thin, elongate ventrolateral sclerites present intersegmentally between pro- and mesothorax and between meso- and metathorax; thoracic legs short, stout, similar in size, shape, and setation ( Fig. 13 View FIGURES 10 – 15 ). Abdomen slightly enlarged, segments not conspicuously depressed or compressed ( Fig. 20 View FIGURES 18 – 27 ); all abdominal segments with sclerotized tergites: segment I with large tergite and a pair of small lateral sclerites, the latter with 3 setae each; segment II with large tergite and a pair of small lateral sclerites, the latter with 2 setae each; segments III–VII each with wide, short tergite, without distinct lateral sclerites, but with 2 lateral setae; segments VIII and IX heavily sclerotized, platelike bearing short and long setae on lateral and posterior margins and short setae on dorsal surface ( Fig. 14 View FIGURES 10 – 15 ), together forming almost circular operculum closing posterior opening of larval case; anal proleg short, setose with pair of long posterior setae; anal claw stout, curved to approximately 90°, with basal peglike setae ( Fig. 15 View FIGURES 10 – 15 ).
Larval case. Length 2.3–3.7 mm (n=14). Made entirely of silk with bits of mineral material incorporated, generally with filamentous algae attached on surface ( Figs. 18, 19 View FIGURES 18 – 27 ); slightly depressed dorsoventrally, with poorly closed dorsal seam; anterior and posterior openings circular.
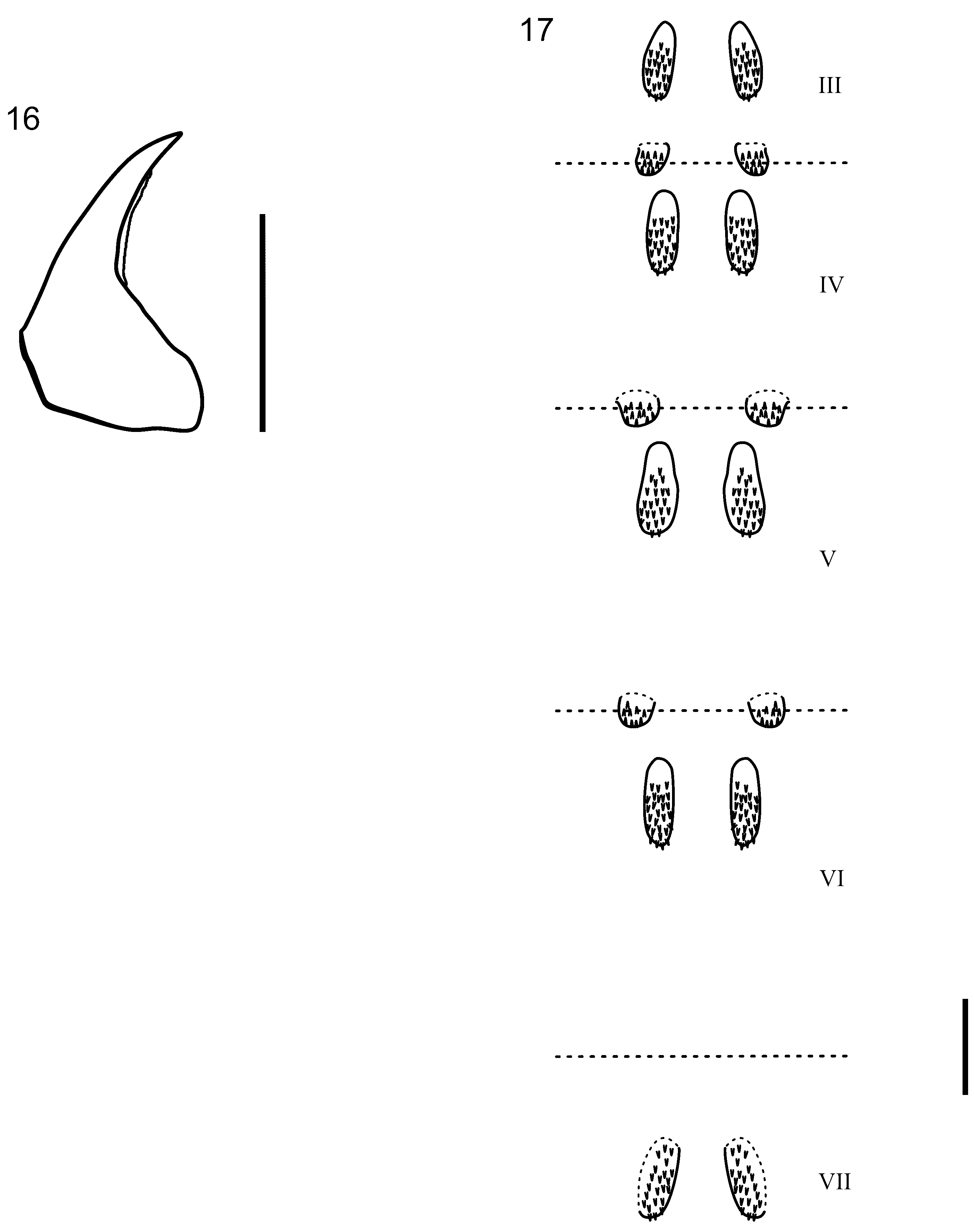
Pupa. Length 2.4–3.0 mm (n=12). General color pale yellow. Mandibles symmetrical, without teeth, but with serrated cutting edge ( Fig. 16 View FIGURES 16 – 17 ). Abdominal segments III–VII with dorsal hook plates ( Fig. 17 View FIGURES 16 – 17 ): segments III–V with 2 pairs of dorsal hook plates, anterior pair elongate oval with anteriorly directed hooks, posterior pair smaller with posteriorly directed hooks; segments VI and VII with only anterior pair of dorsal hook plates present. Terminal abdominal segment simple, without appendages or modifications.
Pupal case. Length 3.0– 3.8 mm (n=12). Anterior end (corresponding to posterior end of larval case) with short ventral stalk attaching pupal case to substrate. Anterior margin truncate; posterior margin rounded, also attached to substrate. Filamentous algae on surface of cases absent or represented by few filaments. Pupal case laterally compressed, in contrast to larval case, but while larva arranged dorsoventrally in case, pupa laterally arranged ( Fig. 21 View FIGURES 18 – 27 ).
Material examined. Holotype male. BRAZIL, Rio de Janeiro State, Rio de Janeiro, Floresta da Tijuca: Parque Nacional da Tijuca, Rio Humaitá, 22°57’30.1”S 43°17’21.4”W, 475 m, 24.x.2009, A.P.M. Santos leg. ( DZRJ).
Paratypes. Same data as holotype, 2 males, 2 females ( DZRJ / MZSP).
Additional material examined. Same data as holotype, 9 larvae, 6 pupae ( DZRJ); same data except, 14.i.2009, A.P.M. Santos, L.L. Dumas, and R.S.G.G. Carrijo, 3 larvae, 3 pupae, 2 pharate males, 2 pharate females ( DZRJ); same data, except, 28.v.2009, A.P.M. Santos and L.L. Dumas, 4 pharate adults, 3 pupae, 2 larvae ( MZSP).
Etymology. The species name refers to the city of Rio de Janeiro, where the species was collected. Carioca is a gentile adjective for the inhabitants of this city. The word “ carioca ” comes from the Tupi language and means “white man’s house”.
Distribution. Rio de Janeiro State, Brazil.
Biology and habitat. Parque Nacional da Tijuca is located in a mountainous area in the center of Rio de Janeiro city, and includes reforested fragments of the Atlantic Forest. The specimens were collected from a second order portion of the Rio Humaitá within the conservation unit. In this portion, the stream was fastflowing and ran over bedrock, in an area exposed to light.
Larvae and pupae were found on rock surfaces, particularly in areas with little water; occasionally they were above the water line. Pupae occurred in small groups and pupal cases were attached to the substrate by the anterior stalks. Gut contents of larvae (n=4) contained almost exclusively diatom frustules, as described for other species ( Holzenthal & Harris 1992, Harris & Holzenthal 1994).
Adults of B. carioca sp. nov. were collected only during the day and, as typical for the genus, ran actively on the rock surface. Some adults of the new species had the uncommon behavior of rotating the body around the axis of the head, after running rapidly and stopping quickly.

| MZSP |
Sao Paulo, Museu de Zoologia da Universidade de Sao Paulo |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Byrsopteryx Flint, 1981
| Santos, Allan Paulo Moreira & Nessimian, Jorge Luiz 2010 |
Byrsopteryx
| Bowles 1999: 45 |
| Flint 1981: 27 |