
Leptobrachella itiokai, Eto & Matsui & Nishikawa, 2016
|
publication ID |
https://doi.org/ 10.5281/zenodo.4504328 |
|
publication LSID |
lsid:zoobank.org:pub:A3E45D0C-26C7-4DF4-B5BC-5C9CDF3BE10B |
|
persistent identifier |
https://treatment.plazi.org/id/F92256D6-0EFA-4B43-BB10-4EA52631DAB3 |
|
taxon LSID |
lsid:zoobank.org:act:F92256D6-0EFA-4B43-BB10-4EA52631DAB3 |
|
treatment provided by |
Carolina |
|
scientific name |
Leptobrachella itiokai |
| status |
sp. nov. |
Leptobrachella itiokai View in CoL , new species
( Figs 3 View Fig , 4 View Fig , 7 View Fig )
Leptobrachella brevicrus: Oberhummer et al., 2014: 66 View in CoL View Cited Treatment ; Eto et al., 2015: 131 (part).
Etymology. The specific name is dedicated to Dr. Takao Itioka of Kyoto University, Japan, who is an entomologist and a core member of the biological researcher consortium in Sarawak.
Material examined. Holotype: KUHE 55895 View Materials , an adult male from Sungai Tapin on Gunung Mulu National Park , Sarawak, Malaysia (Borneo) (04°02’19”N, 114°55’04”E, 1,445 m asl), collected on 23 August 2015 by K. Eto. GoogleMaps
Paratypes: KUHE 55845 View Materials , 55846 View Materials , 55897 View Materials , and SRC 00335 (former KUHE 55898 View Materials ), four males collected from the type locality on 22–24 August 2015 by K. Eto, K. Nishikawa, and Y. A. Rohman .
Diagnosis. A small form of the genus (males 15.2–16.7 mm in SVL); tan or pale-brown dorsally with dark brown markings, V-shape on head, W-shape on shoulder, and triangle- or inverted V-shape on sacral region; tibiotarsal articulation of adpressed limb reaching or beyond eye; tip of digits sharply pointed, forming arrow-like projection. The new species can be distinguished from all other congeners by the combination of the following characteristics: ventrolateral glands small, rounded, usually not in series; side of body with small black blotches; ventrum entirely dark-coloured with tiny light spots; male with thin, thread-like lineae masculinae, although often indistinct in preservative; toe webs poorly developed; chirpy-call ca. 10 kHz in dominant frequency.
Skin dorsally nearly smooth ( Fig. 3A View Fig ); a low supratympanic ridge from eye to axilla; a relatively large tubercle (or gland) on just behind of jaw commissure; sides relatively granular with scattered tubercles/glands ( Fig. 3B View Fig ); chest and abdomen smooth ( Fig. 3C View Fig ); pectoral gland present but small; supraaxillary gland small and rounded; lateral glands also small and rounded; a spaced ventrolateral series of glands present in left side, absent in right side; femoral gland small and indistinct; two and three femoral glands recognised on left and right thigh, respectively.
Description of holotype (measurements in mm). Snoutvent length (SVL) 16.7; habitus moderately slender ( Fig. 3 View Fig A–C); head slightly longer (HL 6.2) than broad (HW 6.0); snout rounded, projecting beyond lower jaw; eye large, length (EL 2.6) slightly greater than snout length (SL 2.2); canthus distinct, slightly constricted; lore slightly oblique, concave; nostril below canthus, about midway between tip of snout and anterior tip of upper eyelid (S-NL 1.1; N-EL 1.1), slightly closer to snout than to canthus; internarial distance (IND 2.1) wider than interorbital distance (IOD 1.8) and upper eyelid (UEW 1.6); pupil diamond shaped; pineal spot invisible; tympanum distinct, diameter (TD 0.9) one-third that of eye, and separated from eye by half of tympanum diameter (T-EL 0.4); vomerine teeth absent; tongue weakly notched posteriorly; subgular vocal sac bipartite with a small opening each near jaw commissure.
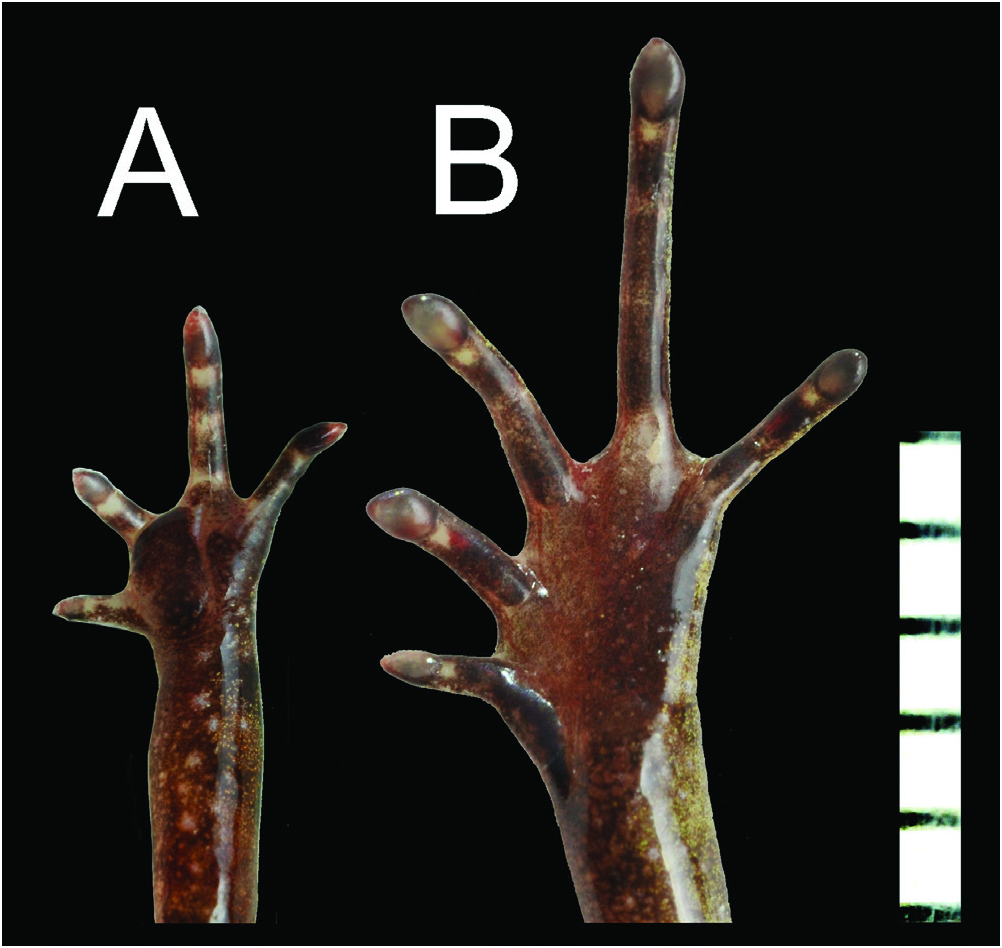
Forelimb slender (LAL 6.7; HAL 3.6); fingers slender, unwebbed; finger length formula: I <II <IV <III ( Fig. 4A View Fig ); finger tips sharply pointed, forming arrow-like projections; no fringes of skin along fingers; inner palmar tubercle large (IPTL 1.5); subarticular tubercles indistinct, replaced by indistinct, low callous tissue; nuptial pads absent. Hindlimb moderately short (HLL 23.8); tibia moderately short (TL 8.1), heels slightly overlapping when limbs held at right angles to body; tibiotarsal articulation of adpressed limb reaching eye; foot (FL 7.2) shorter than tibia; toe length formula I <II <V <III <IV ( Fig. 4B View Fig ); toe tips similar to those of fingers; webbing confined to bases of toes, formula: I 13 / 4 –2 + II 2–3 + III 22 / 3 –4 IV 34 / 5 – 24 / 5 V; toes without lateral fringes; subarticular tubercles obscure but elongate, replaced by low callous tissue; inner metatarsal tubercle low, length (IMTL 1.2) nearly same but slightly shorter than first toe (1TOEL 1.3); no outer metatarsal tubercle.
Colour. In life, dorsally tan on head and body with dark markings, V-shaped marking on upper eyelid and interorbital space, W-shaped on shoulder, and triangle-shaped on sacral region; dorsolateral line light and indistinct; groin with dark marking restricted to cloacal region; side of head darker than dorsum; dark bands below canthus rostralis and supratympanic fold, and upper part of tympanum; a light brown stripe from between nostrils to anteriormost edge of upper jaw; lips irregularly barred with dark brown and white; flank pale-brown with small black blotches; pectoral, supraaxillary, and ventrolateral glands white; darkly pigmented ventrally on head, body, and limbs, except for vocal sac region; trunk darkly pigmented with dense tiny white spots; whitish thread-like lineae masculinae visible through ventral skin; forelimb brown dorsally with dark crossbars on lower arm and fingers; hindlimb brown dorsally with dark crossbars; ventral part of hindlimb dark with tiny white spots except for base of thigh; femoral gland pale, indistinct; iris gold with black reticulation and outer margin of pupil orange.
In preservative, colour and pattern generally faded but not obviously changed except for iris; lineae masculinae became greyish pink and hardly discernible.
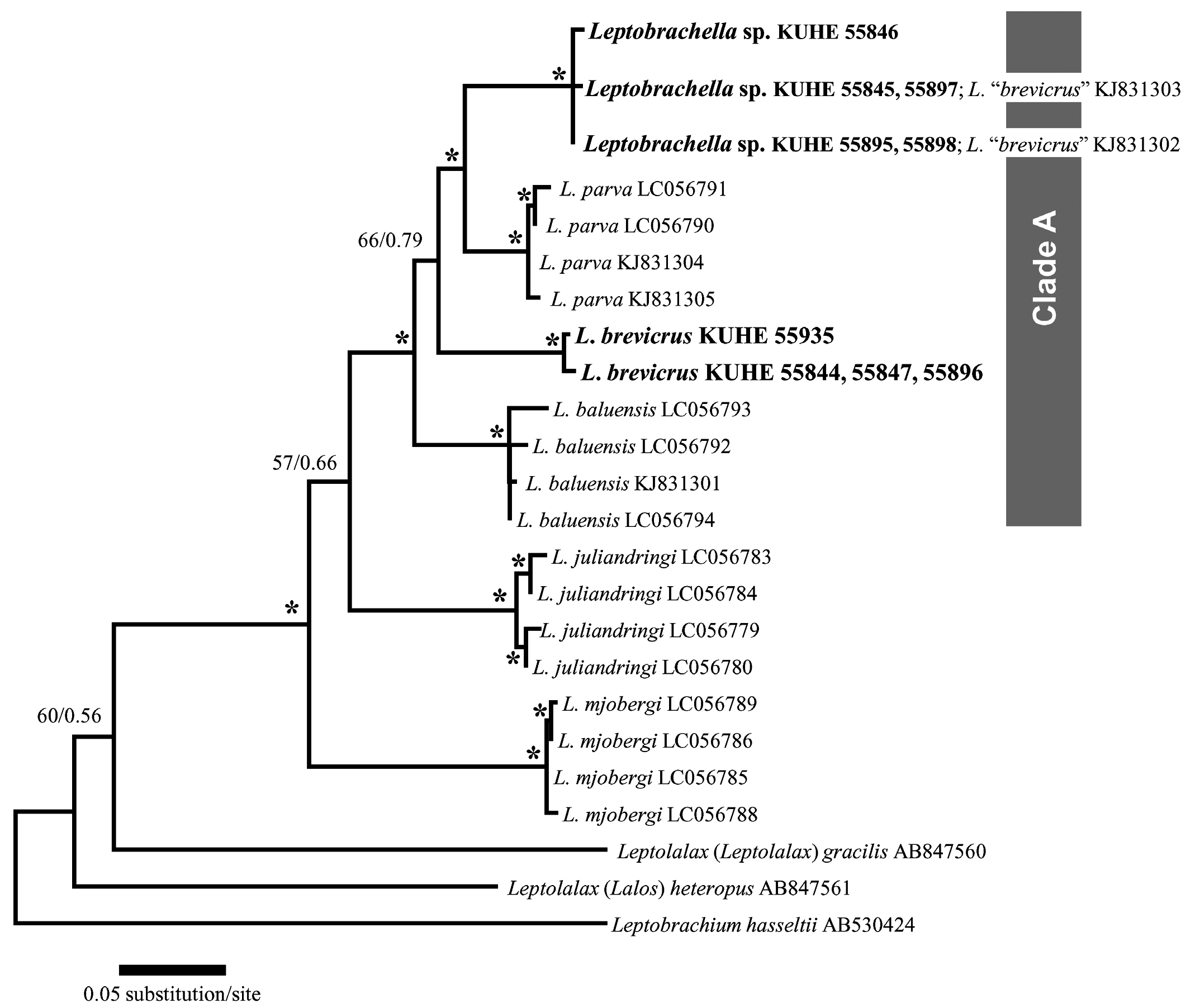
Egg and larva. We could not examine any egg or larva from the type locality, but the tadpole previously reported as that of L. brevicrus from Sungai Tapin (ZMH A09365; Oberhummer et al., 2014) would be assigned to L. itiokai , because it was genealogically embedded in the new species ( Fig. 2 View Fig ; Table 1). It has a morphology typical of the genus Leptobrachella and was found in gravel bed of stream ( Oberhummer et al., 2014; see also discussion).
Variation. Individuals of the type series are generally similar to each other in morphology. Variations in size and body proportions are given in Table 3 together with those of the syntopically occurring L. brevicrus . Tibiotarsal articulation of adpressed limb reaching or extending beyond eye. Dorsal colouration varies among individuals, from tan, yellowish brown to pale brown. Size and number of black blotches on flank variable, but usually small and sparse. Lateral glands not in a series, except for some with an irregular or spaced series on sides of body. Femoral glands two to four in number, although indistinct in some.
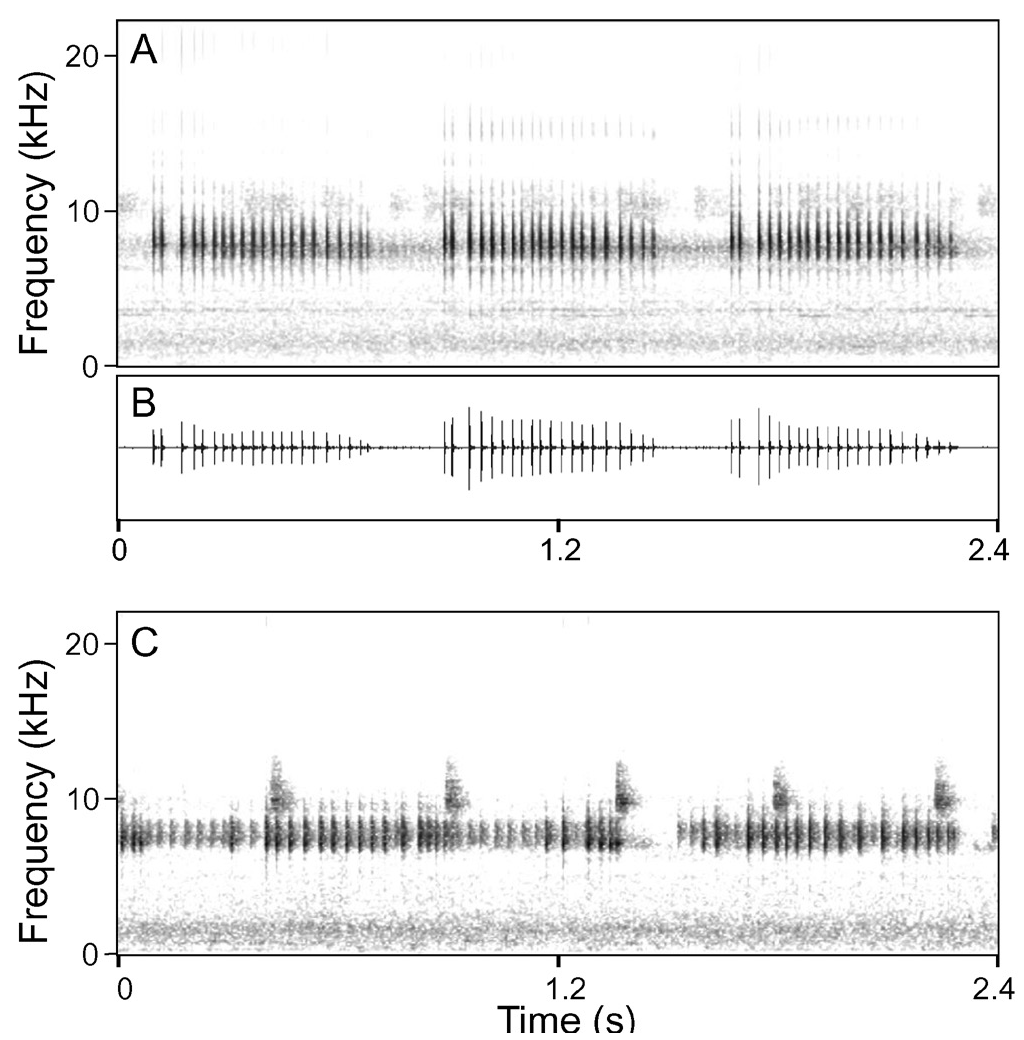
Call characteristics. We examined two calls of the holotype (KUHE 55895) recorded at air temperature of 17.1°C, of which at least one could be the complete call ( Fig. 5 View Fig ). The complete call consisted of a long series of short, chirpy notes
(call duration: 76.5 s; number of notes: 166), and the note repetition rate was 2.2 per s. In the calls, two note types were recognised: notes begin with a discrete pulse (type I: bottom left of Fig. 5 View Fig ), and those without such a pulse (type II: bottom right of Fig. 5 View Fig ). The note type II predominated at the beginning of calls, but was replaced by the type I note from the middle portion to the end of the call. In the note type I, each note lasted 80.1 ± 10.0 (73–106, n = 15) ms and was composed of one discrete and 7–10 subsequent, continuous pulses. On the other hand, note length and number of pulses were 35.3 ± 6.3 (25–51, n = 15) ms and 7–11, respectively, in the type II note. Note interval was 1.07 ± 0.17 (0.84–1.24) s in the beginning and 0.38 ± 0.07 (0.29–0.52) s in the middle and later. The dominant frequency of each note type was essentially same, 10.04 ± 0.03 (9.99–10.08, n = 30) kHz. Multiple harmonics and slight frequency modulation were recognised in both types of notes, and the maximum frequency was over 20 kHz.
Including the holotype, we examined a total of five calls from four individuals recorded at the type locality (air temperatures: 17.1–17.5°C), and obtained ranges of each measurement as follows: note length, 73–106 ms in type I and 21–63 ms in II; number of pulses, 7–10 with one discrete pulse in I and 5–12 in II; note interval, 0.29– 1.24 s, and dominant frequency, 9.98–10.38 kHz.
Comparisons. Leptobrachella itiokai , new species, is similar to L. baluensis , L. brevicrus , and L. parva in appearance. The new species is possibly also similar to Leptobrachella sp. 3 “ baluensis ” from Gn. Mulu reported by Dring (1983) based on literature information. However, the new species can be distinguished from these species, as well as all the other congeners as follows.
By the absence of elongated lateral glands, the new species is differentiated from L. juliandringi (a series of elongate glands present: Eto et al., 2015) and Dring’s (1983) Leptobrachella sp. 3 “ baluensis ” (elongate glands present on posterior part of flanks). The new species is also distinguishable from the other congeners from Borneo and Natuna Islands by the presence of dark pigmentation with small light spots on trunk except for L. baluensis , L. brevicrus , L. parva , and Dring’s (1983) Leptobrachella sp. 3 ( Dring, 1983; this study). Male of the new specie has thin, thread-like lineae masculinae, which is wide, strap-like in L. juliandringi ( Eto et al., 2015) and not evident even on dissection in L. brevicrus and Leptobrachella sp. 3 “ baluensis ” ( Dring, 1983; this study). From L. serasanae Dring, 1983 , the new species is differentiated by the absence of wide black band on flank. From L. palmata , the new species is differentiated by the poor toe webs (toe webs well-developed in L. palmata: Inger & Stuebing, 1992 ). As to body size, the new species (males 15.2–16.7 mm: Table 3) overlaps L. baluensis from Sabah (males 14.9–15.9 mm: this study), L palmata (males 14.4–16.8 mm: Inger & Stuebing, 1992), L. parva (males 14.2–15.1 mm: this study), and Dring’s (1983) Leptobrachella sp. 3 “ baluensis ” (males 15.0–16.0 mm: Dring, 1983), but is smaller than L. brevicrus (males 17.1–17.8 mm: this study) L. juliandringi (males 17.0– 19.1 mm: Eto et al., 2015), L. mjobergi (males 15.7–19.0 mm: Eto et al., 2015), and L. natunae (male 17.6 mm: Günther, 1895). Although only one female specimen is known, L. serasanae would have the body size similar to the new species (female 16.9 mm: Dring, 1983). In the new species, relative length of tibia (RTL = 48.1–52.9%: Table 3) overlaps L. brevicrus (48.2–49.6%), L. natunae (46.1–49.4%: Dring, 1983), and Dring’s (1983) Leptobrachella sp. 3 “ baluensis ” (50.0–52.7%: Dring, 1983), but is shorter than L. baluensis (54.2–56.9%: this study), L. juliandringi (50.0–58.1%: Eto et al., 2015), L. mjobergi (50.1–59.9%: Eto et al., 2015), L. parva (55.9–58.4%: this study), and L. serasanae (59.2%: Dring, 1983).
Other than the morphological traits, L. itiokai has a unique chirpy-call ( Fig. 5 View Fig ), which is distinct form buzz-like advertisement calls of L. brevicrus , L. mjobergi , and L. parva ( Dring, 1983; Eto et al., 2015; Fig. 6 View Fig ). Leptobrachella juliandringi and Dring’s (1983) Leptobrachella sp. 3 “ baluensis ” also have a chirpy-call, but the former has lower (7.00–8.20 kHz at air temperatures of 21.8–23.5°C: Eto et al., 2015), and the latter has higher (12–14 kHz at 22°C: Dring, 1983) dominant frequency, than the new species.
Range. Known only from the type locality, Sungai Tapin on Gn. Mulu , state of Sarawak , east Malaysia.
Natural history. In the type locality, males seem to prefer calling on leaves or branches (ca. 0.2 to 0.6 m above the ground or the water surface: Fig. 7 View Fig ) near steep streams. This habit contrasts to that of syntopic L. brevicrus , which calls in hidden place such as among rocks or dead leaves and at the base of bushes ( Fig. 8 View Fig : Dring, 1983; this study). The oviposition site is unknown but perhaps in gravel portions along streams or water spring, because calling males were often observed near such an environment.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Leptobrachella itiokai
| Eto, Koshiro, Matsui, Masafumi & Nishikawa, Kanto 2016 |
Leptobrachella brevicrus:
| Eto K & Matsui M & Nishikawa K 2015: 131 |
| Oberhummer E & Barten C & Schweizer M & Das I & Haas A & Hertwig ST 2014: 66 |