
Rhacophorus dulitensis
|
publication ID |
https://doi.org/ 10.5281/zenodo.208711 |
|
DOI |
https://doi.org/10.5281/zenodo.5658367 |
|
persistent identifier |
https://treatment.plazi.org/id/E4394C34-D46F-F32E-8196-C225C9863092 |
|
treatment provided by |
Plazi |
|
scientific name |
Rhacophorus dulitensis |
| status |
|
Colour in life (Stage 38, ZMH A13093). The background colour of the body and the tail is ochre or buff brown ( Fig. 1 View FIGURE 1 ). The pigmentation is diffuse and there are melanocytes on the dorsal and lateral body. In dorsal view there is a rhomboidal figure of melanocytes, starting anterior the eyes and reaching the posterior end of the head ( Fig. 1 View FIGURE 1 B) where it transforms into a band posteriorly and terminating at the end of the trunk.
In lateral view, the pigmentation of the trunk slightly decreases from dorsal to ventral and from anterior to posterior. Congregations of larger melanocytes can be found in some specimens in the infraorbital region and may also form a diffuse band from the anteroventral corner of the orbit to the snout, passing ventral to the naris. The epidermal layers covering the abdominal cavity superficially and laterally bear spindle-shaped epidermal melanocytes; the deeper abdominal lining also possesses melanocytes. The dark brown colouration in the lateral trunk region originates from the additive effect of both layers. The lungs are discernible through the skin in dorsal and lateral views as an arched line of spherical shiny shapes dorsal to the gut coil ( Fig. 1 View FIGURE 1 A; Stage 38, ZMH A13093). The spiracular tube is translucent, but clearly visible against the almost black abdominal wall ( Fig. 1 View FIGURE 1 A). Few iridocytes can be present on the spiracular tube and the lateral trunk of some specimens. The hind limbs have a darker pigmentation on the dorsal side. The pigmentation of the muscular part of the tail and of the peripheral fin areas decreases from anterior to posterior, resulting in an almost transparent distal half of the tail. Dorsal and ventral fins are mostly unpigmented. Some specimens show 2–3 circles of assembled melanocytes on the anterior part of the tail, approximately along the horizontal septum of the tail musculature. Melanocytes in the tail are assembled to a line along the vena lateralis, a line at the base of the myosepts of the musculature.
The eyes are black showing scattered golden pigmentation increasingly dense towards the iris and forming a golden ring around the pupil. Golden iridophores are present posteroventral to the eye (region of the cheek). The lateral line system and the nares are of a beige colour. This colour, albeit faded sometimes, is retained in preserved specimens.
In ventral view, the skin of head, trunk, and tail is almost completely unpigmented and translucent. Scattered melanocytes may be present in the gular region and lateral edges of the buccal area. In the latter, some iridocytes can also be present. The red gill tufts and developing forelimbs are visible through the skin in ventral view. The gut coil is clearly visible in ventral view and the gut is dark pigmented ( Fig. 1 View FIGURE 1 C). Iridocytes form a diffuse triangular shaped area that the anterior sector perimeter of the gut coil, right posterior to the heart region ( Fig. 1 View FIGURE 1 C).
External morphological features, shared by Stages 38–39 (ZMH A 10840). The body shape is depressed dorsoventrally, tapering to the snout. In dorsal view, the body contour is ovoid. The oral disc is subterminal in position and cannot be seen in dorsal view. The snout profile is rounded in lateral view with a characteristic elevation dorsal the nares and a bulge just dorsal to the oral disc ( Fig. 1 View FIGURE 1 A). The tail fin rises abruptly at the trunk-tail transition so that the beginning of the tail is clearly marked in lateral view ( Fig. 1 View FIGURE 1 A). The tail contributes 60% of the total length, where earlier larval stages had relatively longer tails than late larval stages in our sample.
The dorsal fin originates in the same level as the tail muscles, at the trunk-tail junction. The ventral fin connects broadly to the trunk. The height of the tail increases gradually, achieving its maximal extent at the level of mid-tail. The dorsal and ventral fin each make up for on third of the maximal tail height (MTH). Posteriorly the tail tapers gradually and terminates in a narrow tip. The muscular part of the tail is moderately high (50% of body depth). The width of the tail’s base equals 27–35% of the maximal trunk width.
The eyes are positioned dorsolaterally. They are placed at 20–26% of the distance between the anterior tip of the snout and the trunk-tail junction. The nares are oriented anterolaterally and are closer to the snout than to the eyes. The internarial distance ( IND) is 40–52% of the interorbital distance (IOD). The spiracle is sinistral. The spiracular tube is directed posteriorly and dorsally (approx. 27° in lateral view, Fig. 1 View FIGURE 1 ), and opens laterally. Its medial part is fused to the abdominal wall (but forms a shallow rim); the lateral circumference of the orifice is free. The opening of the spiraculum is positioned at 60–66% of the head-trunk length. The anal siphon is dextral.
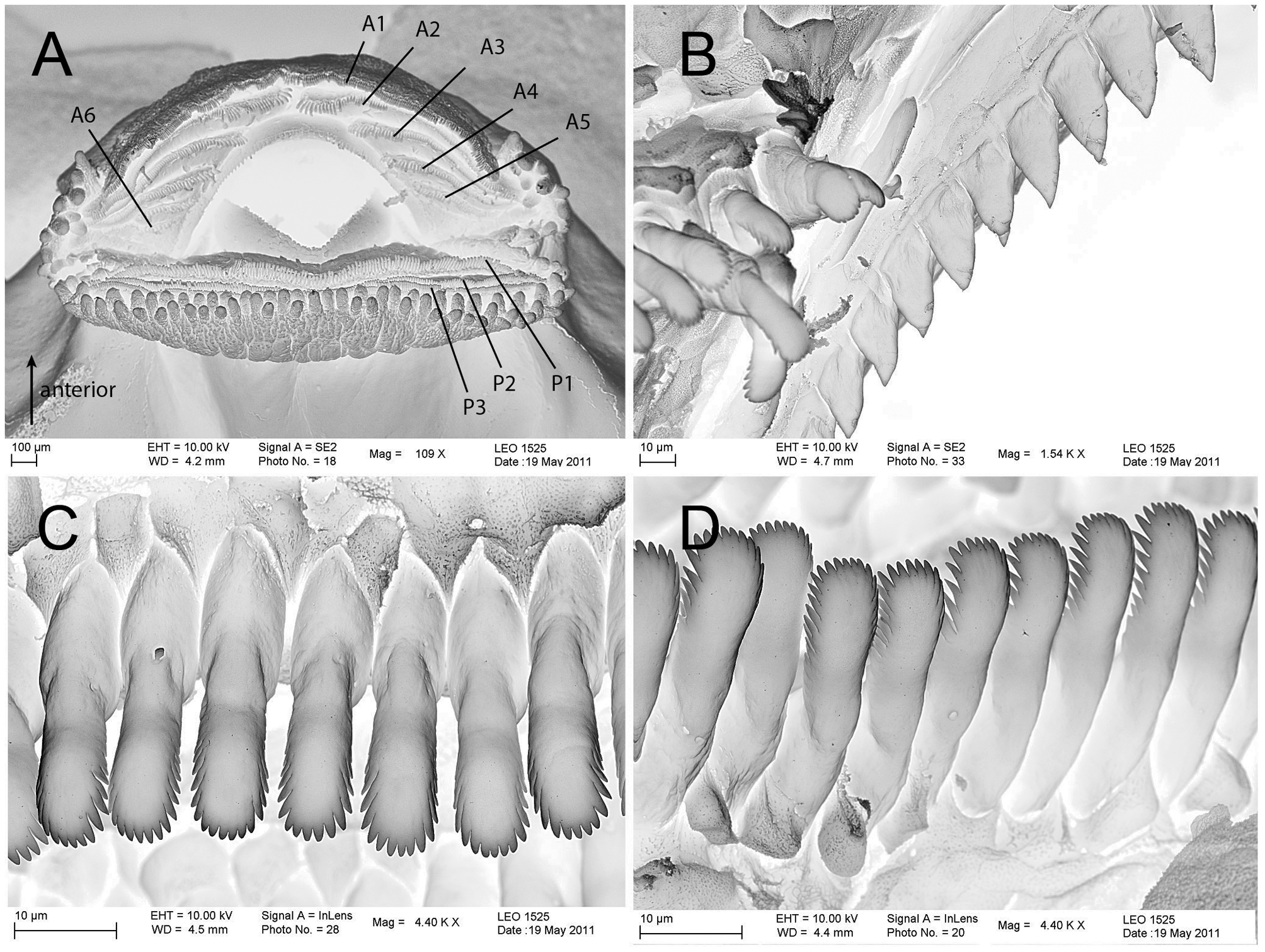
The oral disc is subterminal and can only be seen from the ventral aspect of body. The width of the oral disc is 41–54% of the maximal width of the trunk ( Fig. 1 View FIGURE 1 C). In lateral view and at rest, the lower lip sticks out ventrally at about 90° ( Fig. 1 View FIGURE 1 a). Papillation is present along the margin of the lower lip and lateral parts of the upper lip ( Fig. 5 View FIGURE 5 a). The marginal papillae are arranged mostly bi-serially; papillae are short and rounded.
The labial ridges bear uniserial rows of spoon-shaped keratodonts showing fine incisions along their edges ( Fig. 5 View FIGURE 5 ). The Labial Tooth Row Formula (LTRF) is 5(2–5)/3 to 6(2–6)/3. In the upper lip only keratodont row A-1 is continuous; five divided rows follow caudally. Three undivided keratodont rows are present on the lower lip. The size of the keratodonts decreases in the upper lip from anterior to posterior rows. In the lower lip the size decreases from posterior to anterior. The beaks are well-keratinized, black, and have sharp serrations ( Fig. 5 View FIGURE 5 B). The upper beak is broadly arched in ventral view, whereas the lower jaw sheath is V-shaped.
Variation. In a total of 43 specimens (ZMH A10838, A10837, A13093, A13094; Stages 27 to 41), counts of the number of keratodont rows on the lower lip (P1–P3) were stable, whereas the keratodont row numbers on the upper lip varied: most specimens (47%) had 5(2–5)/3. Another 28% had an additional proximal interrupted row (A6), thus, 6(2–6)/3. Nine specimens (21%) had A6 only on the right or left side of the oral disc. Two specimens had only three (both sides; Stage 33) or four/five anterior rows (left/right; Stage 37). There was no clear correlation between stage and LTRF. Minor differences in pigment patterns were discerned in the samples: the dark band from the eye to the snout was inconspicuous in some ( Fig. 1 View FIGURE 1 A), but more pronounced in other specimen, and the circular assemblages of melanocytes at the base of the tail can be present ( Fig. 1 View FIGURE 1 A) or absent. The density of iridocytes varies individually.
Ecological notes. We found Rhacophorus dulitensis larvae in bodies of standing water. Tadpoles were collected from sun-exposed ponds (50–150 m 2, depth <0.5 m), with muddy substrate. Adult R. dulitensis were encountered at the edges of these ponds. At night, the tadpoles were seen to form schools and to feed at the bottom as well as from surfaces of leaves of macrophytes. Apart from adult R. dulitensis , frogs of the following species were encountered at these ponds: Rhacophorus appendiculatus (Günther, 1858) , R. pardalis Günther, 1858 , Kaloula baleata (Müller, 1836) , Fejervarya limnocharis (Gravenhorst 1829) , Polypedates leucomystax (Boie, 1829) , and Hylarana raniceps (Peters, 1871) .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |