
Gryposuchus jessei Gürich, 1912
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4457.1.9 |
|
publication LSID |
lsid:zoobank.org:pub:C4AF631F-EE68-43CB-88C1-E6586B7511C3 |
|
DOI |
https://doi.org/10.5281/zenodo.5966405 |
|
persistent identifier |
https://treatment.plazi.org/id/E410470D-FFE4-FD5A-BEC0-552EFF5EFB6B |
|
treatment provided by |
Plazi |
|
scientific name |
Gryposuchus jessei Gürich, 1912 |
| status |
|
Gryposuchus jessei Gürich, 1912
( Figure 2 View FIGURE 2 , 3 View FIGURE 3 , 4a and 4b View FIGURE 4 , 5a and 5b View FIGURE 5 )
Holotype: Unnumbered specimen ( Figure 2 View FIGURE 2 ), well preserved distal end of a rostrum, previously housed at Mineralogisch-Geologische Institut (formerly part of the deactivated Naturhistorisches Museum), of Hamburg, Germany ( Gürich 1912). The holotype is considered as lost since the Second World War, probably due to the bombing of 1943 (Ulrich Kotthoffpe, personal communication, 2014).
Revised diagnosis: Gryposuchus jessei presents the following unique combination of features distinguishing it from other species of this genus: 1) the third premaxillary alveoli is aligned with the first maxillary alveoli (shared with G. croizati and G. pachakamue ); 2) the first premaxillary alveoli is bigger than the third one (shared with G. neogaeus and G. pachakamue ); 3) incisive foramen longer than wide, with a medial anterior process within its posterior margin (shared with G. pachakamue ); 4) external naris wider than long (shared with G. colombianus ); 5) well-marked notch between the fourth premaxillary and first maxillary alveoli (shared with G. pachakamue , G. colombianus , and G. croizati ); and, 6) wedge-shaped premaxillary posterior process in dorsal view extending until the fourth maxillary alveoli (shared with G. croizati ).
Occurrence: The holotype was collected on the margins of the Pauynim River (correct spelling Pauini), a tributary of the Purus River, Amazonas State, Brazil ( Gürich 1912; locality F- 2 in Campos et al. 1976; Figure 1 View FIGURE 1 ). The referred material UFAC 1272 was collected in the Sena Madureira locality ( Figure 1 View FIGURE 1 ). Both specimens come from layers regarded to belong to the uppermost sequence of the Solimões Formation (Acre Basin). These layers were dated as upper Miocene ( Latrubesse et al., 2010), based on palynology, vertebrate and invertebrate biostratigraphy from a few localities of the Acre and Amazonas States. It should be noted that the localities of the Gryposuchus specimens studied here were not included in Latrubesse et al. (2010). The Solimões Formation exhibits an extensive temporal range, from the Eocene to Pliocene ( Cunha 2007), and present several reverse faults ( Campos et al. 1976) that can bring older rocks to the surface. Therefore, the lack of proper stratigraphic studies including the Pauini River and the Sena Madureira locality make the age inference dubious (see Souza et al. 2016, for a review of the problems surrounding the age of this stratigraphic unit).
Referred material: Part of a right premaxillary and maxilla ( UFAC 1272 ), housed at the Universidade Federal do Acre, Acre, northern Brazil ( Langston & Gasparini, 1997).
Description: The specimen UFAC 1272 consists of the anterior portion of the rostrum composed by an almost complete right premaxilla, the anteriormost portion of the left premaxilla, including the first premaxillary alveoli, and the anterior portion of a right maxilla ( Figure 3 View FIGURE 3 ). It is about 240 mm long, with the dorsal portion complete and the ventral part formed exclusively by the alveolar border. Both premaxillae present in dorsal view an anterior projection that has the same width as the naris ( Figure 3b View FIGURE 3 and 5b View FIGURE 5 ). Dorsally, a wedge-shaped process of premaxilla reaches the sagittal plane between the region of the third and fourth maxillary alveoli. The contacts between the premaxilla and nasal is not preserved ( Figure 3b View FIGURE 3 ). In ventral view, the incisive foramen is not preserved, and the fourth alveolus is incomplete ( Figure 2a View FIGURE 2 ). The lateral margin of the first maxillary tooth is aligned with the third premaxillary tooth ( Figure 3a View FIGURE 3 and 4b View FIGURE 4 ). Posterior to the notch located at the premaxilla-maxilla suture, the premaxilla becomes thinner giving this bone a wedge-shaped appearance in ventral view ( Figure 3a View FIGURE 3 ). This posterior wedge-shaped process of premaxilla reaches the midline of the skull at the region between the second and third maxillary alveoli ( Figure 3a View FIGURE 3 ). Between the incisive foramen and the second and third premaxillar alveolar border, there are three anteroposteriorly aligned foramina. The alveolar margin is undulated, with marked notches between subsequent alveoli ( Figure 3a and 3b View FIGURE 3 ). There is a well developed lateral expansion at the level of the second premaxillary alveolus ( Figure 3 View FIGURE 3 ). Posterior to the second alveolus, the lateral margins of the premaxilla inclines medially until the region posterior to the fourth premaxillary alveolus. At the premaxillary-maxillary notch ( Figure 3 View FIGURE 3 ), receives the fourth dentary tooth as in other Gryposuchus specimens ( Langston 1973). The maxilla participates only in the posterior end of the premaxillary-maxillary notch.
All four premaxillary alveoli from the right side and only the first from the left side are preserved. The first pair occupies a medial position compared to the others. In terms of size the first and second premaxillary alveoli are the largest with 18 mm and 16 mm of mediolateral diameter, respectively. The third and fourth premaxillary alveoli have 10 mm and 6 mm of mediolateral diameter, respectively. The second premaxillary alveoli is positioned on the laterally expanded portion of the premaxillary. The third and fourth pair of the premaxilla are smaller than the first and second pair, with the fourth being the smallest. The diastema between the first and second premaxillary alveoli is the largest followed by the diastema between the fourth premaxillary and first maxillary alveoli ( Figure 3a View FIGURE 3 ).
Only the right half of the external naris is preserved. This cranial opening is positioned at the level of the second and third premaxillary alveoli ( Figure 3b View FIGURE 3 ). Despite its incompleteness, it is interpreted as having a pentagonal shape, being wider than long, with a transverse, straight anterior margin and a pointed posterior margin ( Figure 3b View FIGURE 3 ). The posterior margin is wider than the anterior margin, because the lateral margins open posterolaterally ( Figure 3b View FIGURE 3 ).
Only a fragmentary portion of the right maxilla is preserved. There are three alveoli preserved, all anteroposteriorly aligned with the third premaxillary alveolus ( Figure 3a View FIGURE 3 ). No significant change in size is observed.
Preserved teeth are found in the first and second premaxillary and in the first maxillary alveoli ( Figure 3a View FIGURE 3 ). Unfortunately, none is complete. The best-preserved one is the first maxillary tooth that lacks the apical region of the crown. This tooth has a conical shape and shows fine parallel striations directed to the apical region ( Figure 3c View FIGURE 3 ).
Remarks: In UFAC 1272 and in the holotype (based mainly on the figures published by Gürich 1912) the external naris, in dorsal view, is pentagonal and anteroposteriorly wider than long, with the anterior margins straight (e.g., Figure 2b View FIGURE 2 and 3b View FIGURE 3 ). The overall outline of the external naris is very similar in both specimens, including the fact that the posterior margin, which corresponds to the apex of the pentagon, is wider than anterior one, due to the posterolateral orientation of the lateral margins ( Figure 2a View FIGURE 2 and 3a View FIGURE 3 ). Furthermore, the third premaxillary alveoli is aligned with the first maxillary alveoli, a feature present in both specimens ( Figure 2a View FIGURE 2 and 3a View FIGURE 3 ) and shared with Gryposuchus croizati and G. pachakamue .
The main differences between UFAC 1272 and the figures of the holotype are found in the ventral portion. The most conspicuous is the presence in UFAC 1272 of three linear aligned foramina that are absent in the figures of the holotype ( Gürich 1912: Figure 2a View FIGURE 2 ). These foramina were also not observed or reported in other Crocodylia specimens, and their significance (as well as their function) remains to be established. The dorsal posterior process of the premaxilla reaches the fourth maxillary alveoli in the holotype, while in UFAC 1272 it reaches the space between the third and fourth maxillary alveoli. All these morphological differences are here treated as individual variations or perhaps a product of the different ontogenetic stages of the specimens (see Mook 1921), with UFAC 1272 representing a younger individual (inferred due to its reduced size in comparison with the holotype). Based on the observations above, UFAC 1272 is regarded as G. jessei , as Langston & Gasparini (1997) have previously suggested.
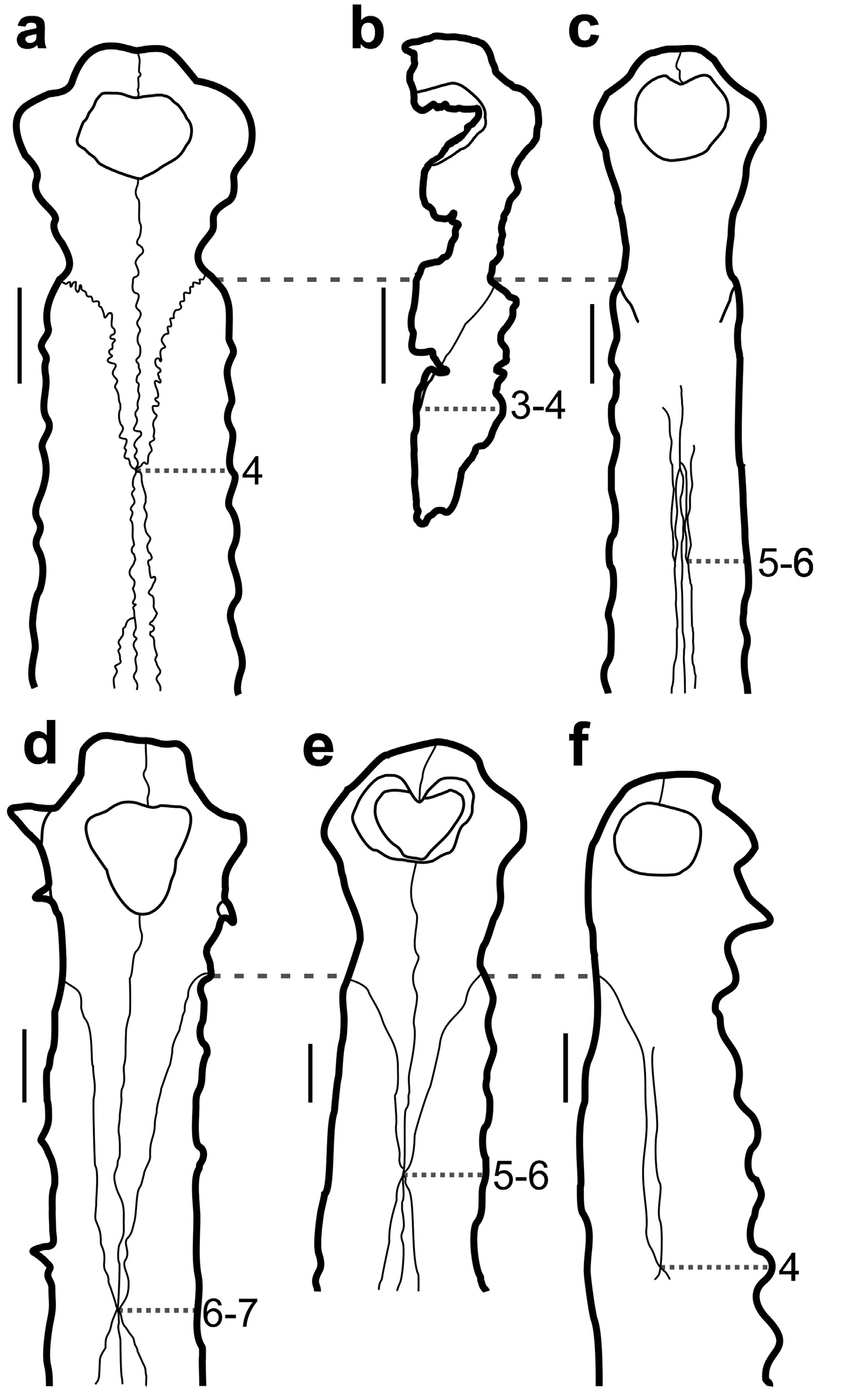
Based on UFAC 1272 and the holotype of Gryposuchus jessei a more detailed comparison with other Gavialoidea species can be provided. Within Gavialoidea, the general shape of the external nares have been used as a phylogenetic character (e.g., character 83 in Brochu 2011, Salas-Gismondi et al. 2016). The features attributed to this character are follows: 1) wider than long; 2) circular; and 3) longer than wide. All anatomical variations described by this character are present in at least one Gryposuchus specimens, being Gr. jessei and Gr. colombianus sharing the condition 1 (wider than long; Figures 4a, 4b and 4e View FIGURE 4 ). Gr. croizati shows condition 2 (circular; Figure 4f View FIGURE 4 ). Gr. neogaeus and Gr. pachakamue present condition 3 (longer than wide; Figures 4c and 4d View FIGURE 4 ). A wider than long external naris (condition 1) is also found in the gavialoids Piscogavialis jugaliperforatus and Eogavialis africanum and an external naris that is longer than wide (condition 3) is found in Ikanogavialis gameroi , Siquisiquesuchus venezuelensis , Gavialis bengawanicus and G. gangeticus .
The external naris of Gryposuchus jessei and Gr. neogaeus present a concave posterior and a rather straight anterior margin, respectively ( Figure 4a, 4b and 4d View FIGURE 4 ), similar to Siquisiquesuchus venezuelensis , Gavialis bengawanicus and G. gangeticus . The species Gr. colombianus and Gr. pachakamue present a medial posterior process on the anterior margin ( Figure 4c and 4e View FIGURE 4 ) similar to Ikanogavialis gameroi , Piscogavialis jugaliperforatus and Eogavialis africanum . Gr. croizati differs from all others by displaying the anterior margin of this opening concave ( Figure 4f View FIGURE 4 ).
Gryposuchus jessei presents the posterior margin wider than the anterior one, which is seen in both UFAC 1272 and in the holotype ( Gürich 1912). This condition differs from all other gavialoids, and could be a unique feature of this species. However, it should be noted that the difference in shape and other morphological details of the external nares have been interpreted as caused by individual variation (e.g., Riff & Aguilera 2008) and sexual dimorphism (e.g., Martin & Bellairs 1977). Therefore, further studies addressing the external naris features are in need to better establish the possible causes of those differences.
The incisive foramen on Gryposuchus species are anteroposteriorly longer than wide as observed in other Gavialoidea, with the exception in G. croizati that has a sub-squared shape ( Riff & Aguilera 2008; Figure 5 View FIGURE 5 ). The species G. jessei (figures of Gürich 1912) and G. pachakamue present a peculiar shape of the incisive foramen, which is triangular and longer than wide, with a small process that invades the posterior margin of this opening, differing from all other gavialoids ( Figures 5a and 5e View FIGURE 5 ).
Although the number of premaxillary alveoli in all Gryposuchus species is four ( Figure 5 View FIGURE 5 ) - a feature also shared by some other gavialoids ( Piscogavialis jugaliperforatus , Ikanogavialis gameroi and Siquisiquesuchus venezuelensis ), there is some variation in size and position. In all Gryposuchus species the second alveoli is the largest and the fourth as the smallest ( Salas-Gismondi et al. 2016). However, size variations between the premaxillary teeth can be observed: 1) the first alveolus is bigger than the third in G. jessei , G. neogaeus and G. pachakamue ( Figure 5a, 5b, 5c and 5d View FIGURE 5 ); 2) the first alveolus is similar in size with the second in G. jessei and G. colombianus (Langston & Gasparini 1998) ; and, 3) the third alveolus of similar size of the second in G. croizati ( Figure 4d View FIGURE 4 ). All specimens referred to Gryposuchus (except one specimen of G. croizati [AMU-CURS-58]) show a marked lateral expansion at the second alveoli, which is followed by a lateral constriction at the premaxilla–maxilla contact, where a diastema is present. There is also a marked reduction in size from the second to the fourth alveolus in all specimens referable to the genus, with the exception of one specimen (AMU-CURS-58) of G. croizati ( Figure 5a, 5b, 5c and 5e View FIGURE 5 ).
The lateral expansion at the level of the second pair (known as Greifaparat sensu Kälin 1933) is present in all Gryposuchus specimens. In G. croizati (specimen MCN-URU- 2002-77), G. jessei , G. neogaeus , G. colombianus and G. pachakamue this structure is well-developed and restricted to the level of the second alveolar pair, being immediately constricted posteriorly to the second alveolar pair until the premaxilla-maxilla suture ( Figure 5 View FIGURE 5 ). However, in the specimen AMU-CURS-58 of G. croizati the lateral expansion is less-developed but is present between second and third alveolar pairs ( Figure 5d View FIGURE 5 ). This expansion creates a straight margin between the second and third alveolar pairs, which progressively becomes thinner posteriorly until reaching the notch on the premaxilla-maxilla suture; this pattern is also observed in Siquisiquesuchus venezuelensis , Piscogavialis jugaliperforatus and Ikanogavialis gameroi .
Gryposuchus jessei , G. pachakamue , G. colombianus and G. croizati present a well-marked notch between the fourth premaxillary and first maxillary alveoli. Also, this notch includes the premaxillary-maxillary suture ( Figure 5 View FIGURE 5 ) and results from a medial compression in this region and a posterior expansion on the maxilla. The other studied Gavialoidea species including G. neogaeus presents only a thinning of the premaxilla posteriorly to the lateral expansion, resulting the alignment of the lateral margins of premaxillae and maxillae ( Figure 5c View FIGURE 5 ).
In all Gryposuchus species and all gavialoids except Gavialis and Hesperogavialis Bocquetin & Buffetaut, 1981 the nasals contact the premaxilla. Two types of contact can be distinguished between these bones: 1) a point contact is observed in Gr. jessei , Gr. neogaeus , Gr. colombianus , Gr. croizati , Piscogavialis jugaliperforatus , and Ikanogavialis gameroi ; 2) an extensive contact with the nasals placed within the premaxillae as observed in Gr. pachakamue ( Figure 4c View FIGURE 4 ) and Eogavialis africanum . Intraspecific variability regarding this contact is common in Gavialoidea. The holotype of Gr. jessei and Gr. Colombianus , for example, have a high asymmetry between the length of the right and left nasals ( Figure 4a and 4e View FIGURE 4 ). Furthermore, in dorsal view, the range of the wedge-shaped posterior process of premaxilla at the premaxillary-nasal contact varies in Gryposuchus . It should be pointed out that due to the kind of contact between premaxilla and nasals in the Gr. pachakamue , the posterior process of the premaxillae (in dorsal view) can be described as “W” shaped ( Figure 4c View FIGURE 4 ). Regarding the distance between the wedge-shaped posterior process of the premaxilla and the maxillary alveoli, Gryposuchus presents three morphotypes: 1) in Gr. jessei and Gr. croizati this process reaches the fourth maxillary alveolus; 2) in G. bengawanicus , G. gangeticus , E. africanum , P. jugaliperforatus , Gr. colombianus and Gr. pachakamue it reaches the space between fifth and sixth alveolus; and, 3) in Gr. neogaeus and I. gameroi it reaches the space between sixth and seventh alveolus. The morphotype observed in Gr. neogaeus could represent an intraspecific variation of the length of this process, and therefore could reflect the same condition observed in Gr. colombianus and Gr. pachakamue .
In Gryposuchus jessei and all other Gryposuchus species the premaxilla, in ventral view, presents a wedgeshaped posterior process, which reaches in length the level of the space between the second and third maxillary alveoli ( Figure 5 View FIGURE 5 ). In contrast, the other studied Gavialoidea this process goes far beyond the fourth maxillary alveoli.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Gryposuchus jessei Gürich, 1912
| Souza, Rafael Gomes De, Riff, Douglas, De Souza-Filho, Jonas P. & Kellner, Alexander W. A. 2018 |
G. pachakamue
| Salas-Gismondi, Flynn, Baby, Tejada-Lara, Claude & Antoine 2016 |
G. croizati
| Riff & Aguilera 2008 |
Gryposuchus jessei
| Gurich 1912 |
Gryposuchus
| Gurich 1912 |
G. jessei
| Gurich 1912 |