
Pterygoplichthys punctatus Günther 1864
|
publication ID |
https://doi.org/10.1590/S1679-62252006000400003 |
|
DOI |
https://doi.org/10.5281/zenodo.5070618 |
|
persistent identifier |
https://treatment.plazi.org/id/E360F967-FFBB-7665-FC22-42B4C2AEE8CC |
|
treatment provided by |
Carolina |
|
scientific name |
Pterygoplichthys punctatus Günther 1864 |
| status |
|
Pterygoplichthys punctatus Günther 1864 View in CoL Fig. 2 View Fig
Material Examined. Brazil, Mato Grosso. AUM 42928, 1, 166.9 mm SL and UF 148742, 2, 78.5-160.1 mm SL. MCP 35755, 1, 203.1 mm SL, Brazil, Mato Grosso, rio Bugre (rio Madeira drainage), about 42 km N of rio Guaporé on highway BR-174, 14°51’35"S, 059°17’57"W, 12 July 2004. MCP 35753, 3, 81.9-117.8 mm SL, creek, tributary of rio Novo which is tributary to rio Guaporé (rio Maderia drainage) on highway BR-174, 14°13’26"S, 59°41’27"W, 12 July 2004. MCP 35754, 2, 156.6- 196.5 mm SL and MNRJ 28496, 1, 167.1 mm SL, creek, tributary of rio Pindaituba, circa 27 km N rio Guaporé on highway BR-174, 14°59’53"S, 59°17’10"W, 12 July 2004. ANSP 180225, 2, 160.0- 183.3 mm SL, MCP 35756, 6, 131.3- 196.5 mm SL, MNRJ 28497, 2, 165.8- 184.1 mm SL, and UF 149606, 2, 154.3- 177.2 mm SL, small river (tributary of the rio Guaporé) on highway BR-174, near Pontes e Lacerda and Comodoro, 14°55’15"S, 59°17’29"W, 12 July 2004. USNM 177716, 1, Brazil, rio Urubu, 1958. NMW 76587, 1 ( Holotype, photographs provided by H. Wellendorf, not measured), rio Guaporé, Amazon system, San Vincente, rio Madeira drainage, Brazil, 8 January 1827, (= Arraial de São Vincente, a village that is no longer extant in northern Mato Grosso; Papavero, 1971 in Weber, 1992). Bolivia. ZMA 109.199, 1, 124.8 mm SL, south of Todos Santos, in pool of standing water, 8 August 1966.
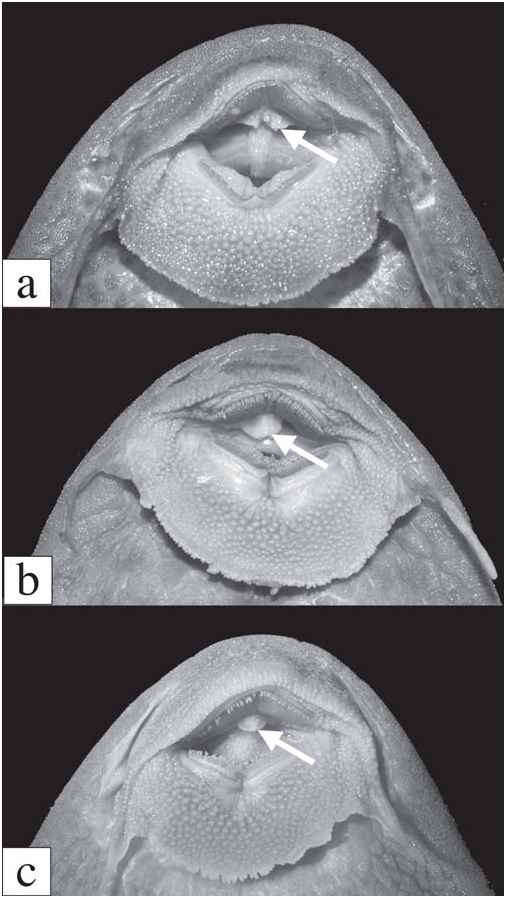
Diagnosis. Pterygoplichthys punctatus can be separated from all other Pterygoplichthys except P. weberi by having the buccal papilla divided medially and lobulate ( vs. entire and simple, Fig. 1 View Fig , or lobulate but not divided medially). Pterygoplichthys punctatus can be separated from P. weberi by having the buccal papilla deeply divided at all ages ( vs. shallowly divided in adult, more deeply divided in juvenile), by having many spots on the abdomen and the fin bases ( vs. few spots), by having a narrower body (SL/cleithral width 3.6-4.0 vs. 3.3-3.4), and by having the lateral keel odontodes sloped posteriorly ( vs. almost perpendicular to the plates) and shorter than the lateral-line plates ( vs. longer than lateralline plates in adult).
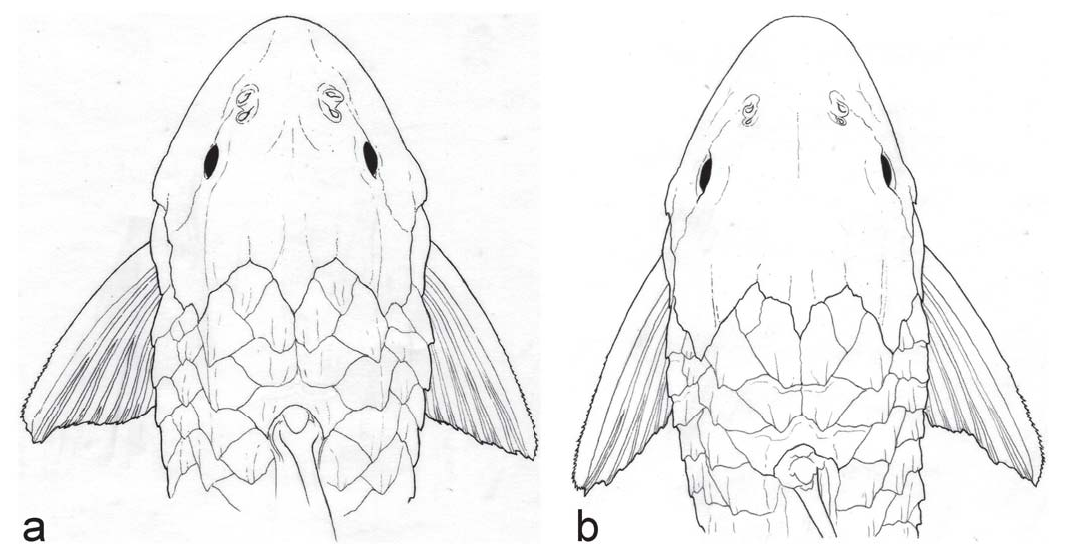
Pterygoplichthys punctatus can be further separated from P. scrophus , P. gibbiceps , P. xinguensis , P. lituratus , and P. parnaibae by lacking an elevated supraoccipital crest ( vs. having an elevated supraoccipital forming an obvious crest higher than the plates in the nuchal region); from P. etentaculatus by having fewer than five hypertrophied odontodes on the cheek plates ( vs. more than five); from P. joselimaianus by having dark spots ( vs. light spots); from P. undecimalis and P. zuliaensis by having a broadly round snout ( vs. a narrow pointed snout, Fig. 3 View Fig ); from P. zuliaensis by having a greater HL to interorbital width ratio (2.3-2.6 vs. 2.0-2.2); from P. multiradiatus by having usually 11 dorsalfin rays (only 3 of 27 with 12 vs. 12-13 fide Weber, 1992); and from P. anisitsi , P. disjunctivus , P. multiradiatus , and P. pardalis by having hypertrophied odontodes on the cheek plates of the adult ( vs. no hypertrophied cheek odontodes).
Description. Member of Hypostominae : Pterygoplichthyini : Pterygoplichthys as diagnosed by Armbruster (2004). Morphometric data given in Table 1 View Table 1 . Largest specimen 205.1 mm SL. Head forming arch from tip of snout to anterior margin of supraoccipital. Supraoccipital and nuchal region forming hump. Body depth decreases from origin of dorsal fin to dorsal procurrent caudal spines, and then increases to caudal fin. Ridge from anterodorsal corner of orbit runs ventral to nares, ending slightly anteroventral of anterior nare. Crest continuing posteriorly from supraorbital ridge with portion on sphenotic and pterotic bent strongly dorsally, posterior portion on pterotic angled dorsally, continuous with keel of mid-dorsal plate series. Dorsal rim of orbit slightly higher than interorbital space.
Keels on lateral plates prominent, formed of ridges of bone and slightly hypertrophied odontodes. Keels present on all plate rows. One or two plates forming short, accessory row between dorsal and mid-dorsal plate series near head; keel on accessory plate row contiguous with but widely separated from keel on dorsal plate series beginning posteriorly to origin of dorsal fin. Keel on anterior plates of dorsal series beginning at midline just anteriorly to posterior margin of supraoccipital, angled to lateral edge of nuchal plate. Ridge on pterotic contiguous with keel on anterior three plates of middorsal plate series; keel on remaining plates of mid-dorsal plate series ventral to that of keel on anterior three plates.
Nares separated by short flap of skin held erect in life. Dorsal, mid-dorsal, median and mid-ventral plate rows complete from head to caudal fin. Ventral plate row begins posteriorly to insertion of pelvic fin and continues to caudal fin. Base of caudal fin covered in elongate, roughly triangular plates. Ventral surface of body (except region dorsal to pectoral fin and ventral to mid-ventral plate row) covered in small plates. Plates on abdomen increase in number with standard length. Head covered in small plates. Frontal, nasal, sphenotic, infraorbitals, pterotic-supracleithrum, suprapreopercle, and supraoccipital supporting odontodes. Opercle with small patch of odontodes in juvenile, none in adult. Platelets covering anteroventral corner of opercle separated from opercle; platelets may be everted to approximately 90° from head. Evertible cheek plates with zero to three slightly hypertrophied odontodes (largest extending approximately to posterior end of opercle).
Dorsal fin long and low in adult, higher in juvenile, consisting of small, V-shaped spinelet, fairly strong spine, usually 11 dorsal-fin rays (three of 27 with 12); adpressed dorsal fin does not reach adipose fin. Caudal fin forked, lower lobe longer than upper. Pectoral-fin spine strong, reaches posteriorly to pelvic-fin rays when depressed ventral to pelvic fin; cleithrum with exposed process dorsal to pectoral-fin rays and strongly angled dorsally; pectoral fin inserted on same plane as pelvic fin such that spine, when depressed parallel with body, lies on top of and in contact with pelvic fin. Pelvicfin spine thin, flexible, reaches barely to base of anal fin. Anal fin with relatively strong, unbranched first ray supporting odontodes. Adipose fin consisting of single median preadipose plate and strong, pointed spine; adipose-fin membrane not reaching procurrent caudal-fin spines. Dorsal fin II,11, pectoral fin I,6, pelvic fin I,5, anal fin I,4, caudal fin usually I,14,I (one specimen I,13,I). Jaws weakly angled, dentaries forming angle of approximately 90º. Teeth bicuspid, median cusp short, lateral cusp about half length of median cusp, stalk moderately long; 20-36 dentary teeth, 19-38 premaxillary teeth. Median plates 26-29 (mode 28), adipose-caudal plates four to six (mode five), anal-caudal plates 11-13 (mode 12), dorsal-adipose plates five to seven (mode six). Buccal papilla lobulate and divided medially ( Fig. 1a View Fig ).
Color in alcohol. Body and fins light brown to gray and covered with black spots. Spots smallest on snout, progressively larger to caudal peduncle; spots on fins nearly as large as eye. Venter covered with discrete (uncoalesced) spots; spots smallest on throat and becoming progressively larger to interpelvic area. Spots proportionally larger and more distinct on juvenile. Caudal peduncle lighter ventrally than dorsally.
Range. Known from the rio Madeira drainage and the rio Urubu of Brazil ( Fig. 4 View Fig ). Weber (1992) additionally notes localities in the rio Purus and the rio Tocantins basin; however, we did not confirm these identifications. Also, not all specimens available to Weber (1991, 1992) from the upper Amazon were available to us, and their identity needs to be verified.


| AUM |
Auburn University Museum of Natural History |
| UF |
Florida Museum of Natural History- Zoology, Paleontology and Paleobotany |
| MCP |
Pontificia Universidade Catolica do Rio Grande do Sul |
| MNRJ |
Museu Nacional/Universidade Federal de Rio de Janeiro |
| ANSP |
Academy of Natural Sciences of Philadelphia |
| USNM |
Smithsonian Institution, National Museum of Natural History |
| NMW |
Naturhistorisches Museum, Wien |
| ZMA |
Universiteit van Amsterdam, Zoologisch Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |