
Oligodon arenarius, Vassilieva, Anna B., 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4058.2.4 |
|
publication LSID |
lsid:zoobank.org:pub:1FDCAFAB-1E21-43A7-9543-E9071EA48FA7 |
|
DOI |
https://doi.org/10.5281/zenodo.6103018 |
|
persistent identifier |
https://treatment.plazi.org/id/E3129514-090D-FF97-4093-FB24FBBEFB0A |
|
treatment provided by |
Plazi |
|
scientific name |
Oligodon arenarius |
| status |
sp. nov. |
Oligodon arenarius sp. nov.
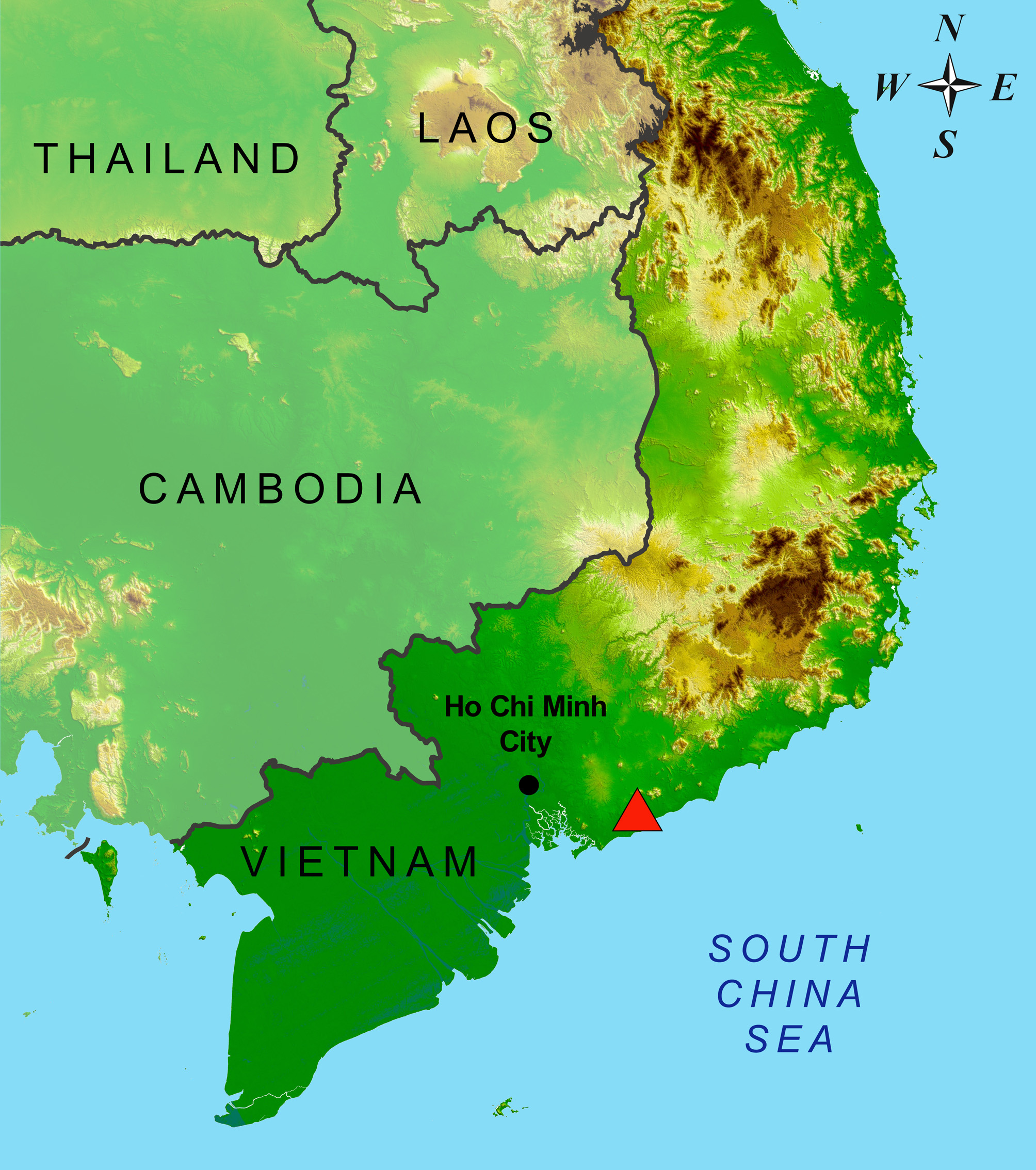
Holotype. ZMMU R- 14503 (field ID ZMMU ABV –00813), adult male from Binh Chau–Phuoc Buu Nature Reserve, Xuen Moc District, Ba Ria–Vung Tau Province, southern Vietnam ( Fig. 1 View FIGURE 1 ), coordinates 10°29'46"N, 107°27'54"E, elevation 5 m a.s.l., collected by Anna B. Vassilieva on 14 November 2014.
Paratypes. ZMMU R- 14002 (field ID ZMMU NAP- 03137), adult male from type locality: Binh Chau–Phuoc Buu Nature Reserve, Xuen Moc Disctrict, Ba Ria–Vung Tau Province, southern Vietnam (10°32'18"N, 107°28'37"E, 43 m a.s.l.), collected on 8 November 2010 by Anna B. Vassilieva; ZMMU R- 14002 (field ID ZMMU NAP- 03138), adult female from type locality (10°29'50"N, 107°27'58"E, 5 m a.s.l.) collected on 9 November 2010 by Anna B. Vassilieva; VNMN 0 4724 (field ID NAP- 03884), adult female from type locality, environs of the resort Ho Coc (10°29'54"N, 107°28'08"E; 8 m a.s.l.), captured on 10 July 2012 by local people and collected by Anna B. Vassilieva and Nikolay Poyarkov; ZMMU R- 14504, adult female from type locality (10°29'41"N, 107°27'46"E; 7 m a.s.l.), collected on 16 November 2014 by Anna B. Vassilieva.
Diagnosis. The species is allocated to the genus Oligodon based on the following characters considered to be diagnostic for the genus ( Smith, 1943): posterior maxillary teeth strongly enlarged and compressed; palatine teeth well developed or vestigial; head short, not distinct from neck, usually displaying a specific coloration pattern; head scalation complete or reduced; eye moderate, with rounded pupil; rostral scale enlarged, extending on to the upper surface of the snout and partly separating the internasals; body cylindrical; scales smooth; ventrals rounded or obtusely keeled laterally; subcaudals paired.
The new species is distinguishable from other species of the genus by a combination of the following features: (1) medium size in adults (TL 336–389 mm); (2) 17(19)–17–15 dorsal scale rows; (3) 6–8 maxillary teeth, the posterior three being enlarged; (4) head scalation lacking a loreal but including a presubocular (except one specimen), (5) nasal divided; (6) two postoculars; (7) 131 – 144 ventrals; (8) 36–60 subcaudals (36–40 in females, 58–60 in males); (9) anal plate entire; (10) tail quite long for the genus in males (TaL/TL = 0.26–0.28) and moderate in females (TaL/TL = 0.13–0.17); (11) unforked hemipenes without spines; (12) head coloration pattern including ocular band, temporal bands and chevron-shaped mark on nape; (13) dorsal coloration without specific pattern, uniform or with dark speckling; (14) ventrals pinkish in life, immaculate; (15) iris bicolored with silvery upper third and dark brown lower two thirds.
Description of holotype. Main morphometric characters are given in Table 1.
Body ( Fig. 2 View FIGURE 2 ) cylindrical, rather robust, tail long for the genus (TaL/TL = 0.26), slender, with acuminated tip; shallow artificial groove caused by the temporary (removed) ligature is seen at the base of tail; head ( Fig. 3 View FIGURE 3 ) short (HL/SVL = 0.04), ovoid in dorsal view, somewhat triangular in profile view, faintly distinct from neck, somewhat depressed dorsoventrally; snout narrowed, triangle with blunt tip; snout tip strongly extending beyond the lower jaw; large oval nostril piercing the central part of nasal; eye rather small, approximately 0.17 times the head length; pupil rounded.
Dentition. Maxillary teeth: 8 functional teeth on the right and 7 on the left side, the posterior three teeth being strongly enlarged, slightly curved posteriorly, dagger-shaped; no distinct gap before the enlarged teeth. Palatine dentition well developed.
Body scalation. Dorsal scale rows 17 - 17 - 15, all smooth; outermost dorsal scale row slightly enlarged; 134 ventrals, obtusely keeled laterally; preventrals absent; anal plate entire; 58 subcaudals, all paired; terminal caudal scale forming an acuminated tail cap.
Head scalation. In lateral view ( Fig. 3 View FIGURE 3 A), head scalation comprising a subrectangular nasal, pierced by large nostril and divided almost vertically by a prominent suture running through the posterior half of nostril; single rectangular preocular, slightly higher than wide, separated from nasal by the lateral part of the prefrontal; single small subrectangular presubocular; two small postoculars, almost equal in size; eight supralabials: dorsally, I in contact with nasal, II in contact with nasal and prefrontal, III in contact with prefrontal and presubocular, IV in contact with presubocular and eye, V in contact with eye, VI in contact with lower postocular and anterior temporal, the largest VII in contact with anterior temporal, VIII in contact with lower posterior temporal; 1+2 temporals, the anterior one subrectangular, elongated and narrow, the upper posterior smoothly pentagonal, approximately equal to the anterior one, elongated, the lower posterior the smallest, rhomboid. In dorsal view ( Fig. 2 View FIGURE 2 B), head scalation comprising a rostral, two internasals, two prefrontals, two supraoculars, a frontal, and two parietals. Rostral large, wider than high, extending on to the dorsal surface of the snout and inserting deeply between internasals. Internasals wider than long, narrowing and slightly curving back towards the median suture. Prefrontals large, pentagonal, approximately 1.5 times wider than internasals, curving down laterally to the loreal area; lateral portions of prefrontals contacting posteriorly with the upper preocular and ventrally with the II – III supralabials. Supraoculars large, elongated, widening posteriorly, approximately half as wide as long. Frontal large, smoothly hexagonal, slightly longer than wide, posterior angle rather acute. Parietals irregularly pentagonal, slightly longer and wider than frontal, bordered laterally by the first and the upper second temporals and posteriorly by four small scales; no enlarged nuchal scales present. In ventral view ( Fig. 2 View FIGURE 2 C), nine infralabials on the left and seven on the right side, III+IV and VII+VIII being fused on the right side; I medially in contact with each other and rostrally with mental, II – V in contact with two enlarged chin shields; mental small, triangular; two enlarged, elongated chin shields, the anterior pair being approximately twice longer than the posterior pair along the midline.
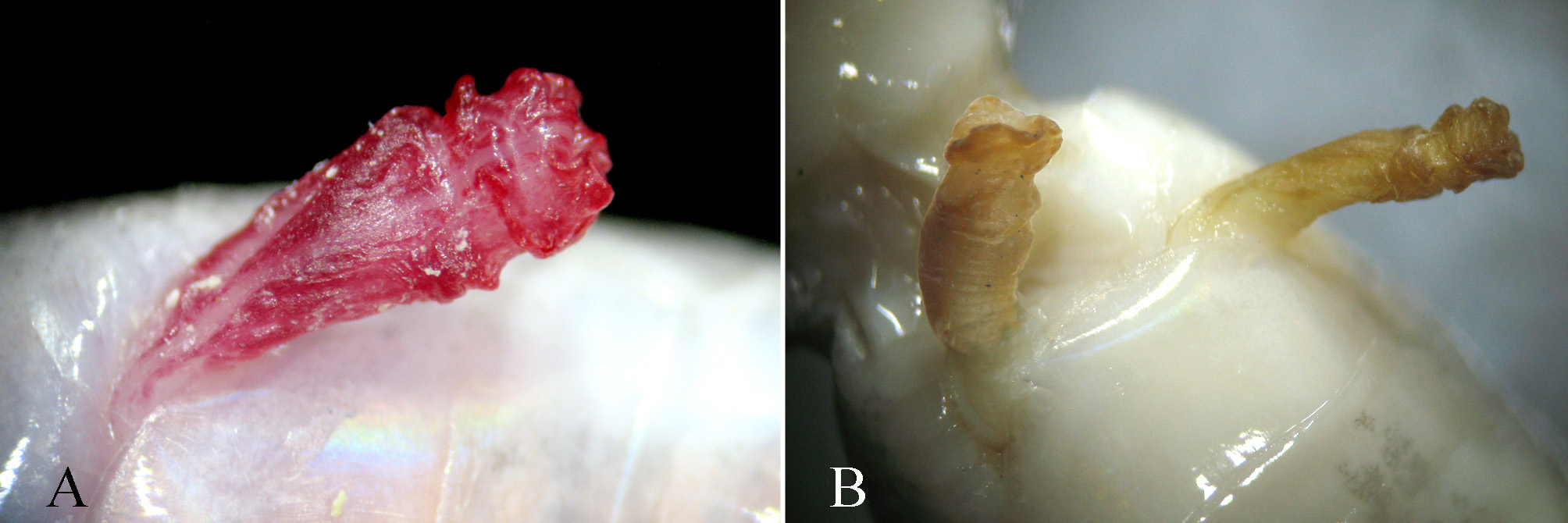
Hemipenis ( Fig. 4 View FIGURE 4 ). Hemipenis rather short, the partly everted organ hardly reaching IV subcaudals; unforked, without spines or obvious papillae, proximal part with smooth surface, distal part with flat and smooth circular folds.
Coloration. In life ( Fig. 5 View FIGURE 5 AC), dorsal coloration pale sandy-gray, with slightly more saturated yellowish tint on 4 – 5 upper dorsal and vertebral scale rows; all dorsal scales scattered with minute dark brown spots. Some dorsal scales (except of three lowerest rows on each side) edged by dark brown forming a diffuse speckled or dashed pattern on dorsum. On nape two pairs of irregular dark spots formed by brown-colored halves of 2 – 3 scales are most visible along the paravertebral zone. Dark speckling less conspicuous on the caudal part of dorsum. On tail, small brownish speckles fuse into two very faint diffuse lines along two adjacent vertebral scales rows edging a slightly paler vertebral line. Ventrals immaculate; gular and anterior ventral scales creamy-white, belly salmonpink with more intensive coloration caudally. Subcaudals salmon-pink in the anterior part of the tail, gradually whitening toward the tail tip. Head ( Fig. 3 View FIGURE 3 AB, 5B) ground coloration sandy-gray with yellowish tint on snout; supralabials slightly paler. Darker head pattern include a faintly visible brown crescent-shaped interocular band partly covering the internasals, prefrontals, supraoculars and frontal scales; the anterior margin of the interocular bar diffuse, the posterior rather clear, edged with dark brown; two diffuse brown markings on the posterior part of each supraocular; short brown oblique subocular stripe extending from the lower margin of each orbit towards the mouth and covering parts of V and VI supralabials; brown oblique temporal bars with sharp, dark-edged anterior end and diffuse posterior one, starting close to the anterior margins of parietals and running down towards the neck just behind the jaw angle; large arrow-shaped chevron, brown with clear darker edges, extending from the center of frontal and medial part of parietals to the occiput and nape, reaching the second paraventral scale row on each side laterally. Iris bicolor, with silvery upper third and dark brown lower two thirds; tongue pinkish-red ( Fig. 5 View FIGURE 5 B).
In preservative ( Fig. 2 View FIGURE 2 AB, 3A – C), all brown and yellow tints fade and turn slightly to gray or cream. Ventral surface yellowish-cream, very faint small grayish spots are visible in the middle of the anterior subcaudals and along the lateral margins of the posterior ventrals.
Description and variation of the paratype series. The most important characters of the four other available specimens, all considered paratypes, are given in Table 1.
A
Specimen ASR MSR PSR SL IL Teeth Nasal Anal PreOc PrSbOc PostOc Temp VEN SC SL contacting eye
14002 17 17 15 7 9 D E 131 60 IV
7/6 1 1 2/1 1+2
14002 19 17 15 8 9 D E 142 40 IV – V
8/6 1 1 2 1+2
VNMN 0 4724 17 17 15 8 9 D E 143 36 IV – V
7/7 1 no 2 1+2
14503 17 17 15 8 8 /9 D E 134 58 IV – V holotype 8/7 1 1 2 1+2
14504 19 17 15 8 9 D E 144 37 IV – V
6/7 1 1 2 1+2
Morphological characters mostly agree with those described for the holotype with few exceptions.
Body proportions. Relative tail length differs significantly in the male paratype (TaL/TL 0.28) and female paratypes (average TaL/TL 0.15, three specimens).
Dentition. 6–8 functional maxillary teeth in all specimens, the posterior three being strongly enlarged and dagger-shaped; palatine dentition well developed.
Body scalation. All scales are smooth, small, ovoid; first and second rows slightly enlarged, the vertebral row does not differ in size from other median dorsal rows; ASR varies 17/19, MSR and PSR are equal in all specimens, 17 and 15, respectively. Ventrals obtusely keeled laterally, 131 in one male (134 in the holotype), 142–144 in three females; anal plate entire, subcaudals paired, 60 in one male (58 in the holotype), 36–40 in females.
Head scalation. Generally as for holotype. In all specimens the nasal is divided through the nostril by a suture; loreal absent, preocular single, presubocular present in three specimens (and in the holotype) and absent on both sides in one specimen (VNMN 04724); two postoculars present with one exception (ZMMU R-14002 male), when the asymmetrical condition is observed (2/1). Eight supralabials (VI and V contacting eye) with one exception (ZMMU R-14002 male), when the IV supralabial (occupying the place of VI and V supralabials of other specimens) is unusually large and the single to contact the eye. Nine infralabials in all paratypes. No variability observed in temporal scales.
Coloration in life. Dorsal background coloration varies from sandy-orange (ZMMU R-14002 male, Fig. 6 View FIGURE 6 A) to orange (ZMMU R-14002 female, Fig. 6 View FIGURE 6 B), yellowish-gray (VNMN 0 4724, Fig. 6 View FIGURE 6 C) or gray (ABV-00814, Fig. 6 View FIGURE 6 D); orange or yellow tint always being more saturated on the dorsum (sometimes forming a faint and diffuse vertebral line, as in female paratype ZMMU R-14504) and gradually turning paler to lower flanks. Blackish or dark brown speckles on dorsum are almost absent (ZMMU R-14002 male) or quite numerous and evident, evenly dispersed (VNMN 04724) or forming two more discrete paravertebral rows (ZMMU R-14504). On neck these dark speckles are always more discrete and may form two short paravertebral streaks (VNMN 04724), on tail they may fuse into faint paravertebral lines (ZMMU R-14504).
Head pattern similar in all specimens, including faintly visible intraocular band and more clear brown-colored temporal bars and chevron. Ventrals and subcaudals immaculate in all specimens, in paratype female ABV-00814 salmon-pink as in holotype; vital belly coloration for other paratypes was not documented.
In preservative, coloration fades slightly, especially the orange and yellow tints, which change to beige. Ventrals and subcaudals creamy-white or ivory.
Distribution. To date known only from the type locality (Binh Chau–Phuoc Buu Nature Reserve).
Habitat and natural history. Four specimens (ZMMU R-14002 female, VNMN 0 4724, ZMMU R-14503, ZMMU R-14504) were collected in the coastal area along the Loc An–Binh Chau secondary road. The landscape comprises gently sloping low sand dunes covered with more or less sparse trees or high bushes ( Fig. 7 View FIGURE 7 A), including Buchanania siamensis (Anacardiaceae) , Casuarina equisetifolia (Casuarinaceae) , Blachiaja trophifolia, Suregada multiflora (Euphorbiaceae) and Randia dasycarpa (Rubiaceae) , with the locally present herbs Pandanus odoratissimus (Pandanaceae) , Vitex rotundifolia (Verbenaceae) , Spinifex littoreus (Poaceae) and Ipomoea pescaprae (Convolvulaceae) . The area's natural character has been severely degraded by human activities, and most of the coastal area is occupied by resorts or private homes.
One specimen (ZMMU R-14002 male) was collected approximately 4 km from the seacoast, on the edge of a lowland forest ( Fig. 7 View FIGURE 7 B), where open sandy areas are present and the co-dominant species are Dipterocarpus chartaceus , Shorea roxburghii , S. siamensis (Dipterocarpaceae) , Melanorrhoea laccifera (Anacardiaceae) and Irvingia malayana (Irvingiaceae) , with Melaleuca cajuputi (Myrtaceae) dominating in seasonally wet areas. The natural character of the area has been profoundly altered by selective logging and cattle pasturing.
All specimens were collected on the ground. They were active during the day (one specimen) or after dark (20:00–22:00, three specimens). In attempts to avoid capture two of the specimens dived into the loose sand and moved swiftly under the sand layer, reappearing on the surface after 1.5– 2 m. The specimen ZMMU R-14504 was found while in the process of swallowing an adult microhylid frog Microhyla pulchra (Hallowell) . The digestive tract of other specimens contained some amounts of sand, implying that they foraged on the ground.
In the Binh Chau–Phuoc Buu Nature Reserve, Oligodon arenarius sp. nov. coexists with other Oligodon species: O. deuvei (data of A. Vassilieva and N. Poyarkov), as well as O. fasciolatus and one unidentified Oligodon sp. ( Nguyen & Hoang 2013).
Etymology. The new species name is an adjective in the nominative case, masculine gender, derived from the Latin word "arena" meaning "sand", "sandy land". The name is intended to reflect the evident preference by the new species for sandy coastal habitats and the snake's remarkable ability to plunge into the sand and move under it.
Comparisons. Oligodon arenarius sp. nov. differs from all other species of the genus by the unique combination of the following characters: 17 DSR, the absence of a loreal scale, unforked hemipenis without spines or obvious papillae, the absence of a specific dorsal coloration pattern (blotches, crossbars or longitudinal lines) and the expressed sexual dimorphism in the relative tail length and number of subcaudals.
The absence of a loreal is a rather rare feature in kukri snakes. In particular, among the 22 Oligodon species known from Indochina, this element of head scalation is consistently absent in only three species: O. annamensis Leviton ( Vietnam, Cambodia), O. catenatus (Blyth) ( Vietnam, Thailand, Myanmar) and O. lacroixi Angel et Bourret ( Vietnam, China); also, two Indochinese species occurring in Vietnam, O. mouhoti (Boulenger) and O. macrurus (Angel) , lack the loreal scale facultatively. Oligodon arenarius sp. nov. differs from them by the following features: from O. annamensis , O. catenatus and O. lacroixi , by a greater DSR count (17 vs. 13, 13 and 15, respectively); usually present presubocular; 8(7) supralabials vs. 6(5), 6 and 5, respectively; 9(8) infralabials vs. 6, 7 and 6, respectively; a lesser number of ventrals (131–144 vs. 148–170, 179–212 and 162–178, respectively) and a dorsal coloration pattern (uniform or with dark speckling vs. thin light crossbars in O. annamensis , chained vertebral line in O. catenatus and four indistinct longitudinal lines in O. lacroixi ); additionally, Oligodon arenarius sp. nov. differs from O. annamensis and O. lacroixi by a greater number of subcaudals (58–60 vs. 44–46 in males, 36–40 vs. 30 in females of O. annamensis , 29–34 in O. lacroixi ) and from O. catenatus and O. lacroixi by an entire anal plate instead of divided one; Oligodon arenarius sp. nov. differs from O. mouhoti by a longer tail in males (0.26–0.28 vs. 0.17–0.19) and a greater number of subcaudals in males (58–60 vs. 39–43), as well as by dorsal coloration features (vertebral stripe and two large dark blotches on tail in O. mouhoti ) and the hemipenial morphology (see below); from O. macrurus , by a relatively shorter tail in males (TaL/TL 0.26–0.28 vs. 0.34–0.37, calculated after the data of Geissler et al. 2011, or 0.31 in one specimen, P. David, pers. comm.), a markedly lesser number of subcaudals in males (58–60 vs. 73–94), a slightly lesser number of ventrals in males (131–134 vs. 139– 145) and the hemipenial morphology (see below).
Oligodon arenarius sp. nov. also differs from many congeners by its hemipenial morphology (unforked hemipenis devoid of spines and papillae). In particular, it differs from all the members of the informal O. taeniatus group (sensu Smith 1943, David et al. 2008b) with Indochinese distribution, namely O. taeniatus (Günther) , O. pseudotaeniatus David, Vogel & Van Rooijen , O. deuvei David, Vogel & Van Rooijen , O. barroni (Smith) , O. moricei David, Vogel & Van Rooijen and O. mouhoti discussed above, which possess deeply forked hemipenes. Additionally, Oligodon arenarius sp. nov. differs from them externally by the dorsal coloration (all members of the O. taeniatus group possessing vertebral stripes or blotches) and by the following characters: from O. taeniatus , by 17 vs. 19 DSR and a greater subcaudals count in males (58–60 vs. 38–48); from O. pseudotaeniatus , by 9 vs. 8 infralabials and a greater subcaudals count in males (58–60 vs. 44–46); from O. deuvei , by a slightly lesser ventrals count (131–134 vs. 140–147 in males, 142–144 vs. 147–155 in females) and a greater subcaudals count in males (58–60 vs. 36–47); from O. barroni , by a relatively longer tail in males (0.26–0.28 vs. 0.17–0.19), a greater subcaudals count in males (58–60 vs. 36–48) and females (36–40 vs. 28–35) and the immaculate belly coloration; from O. moricei , by a lesser ventrals count (131–144 vs. 175), 6–8 maxillary teeth vs. 12 and a usually present presubocular.
Oligodon arenarius sp. nov. also differs by its hemipenial morphology from all the members of the informal Oligodon cyclurus group (sensu Smith 1943, David et al. 2008a, Green et al. 2010). The species of Oligodon cyclurus group are distributed in Indochina, China and India, and possess deeply forked hemipenes without spines but sometimes with papillae; these features were documented for O. cattienensis Vassilieva, Geissler, Galoyan, Poyarkov, Van Devender & Böhme , O. chinensis (Günther) , O. cyclurus (Cantor) , O. fasciolatus (Günther) , O. formosanus (Günther) , O. juglandifer (Wall) , O. kampucheaensis Neang, Grismer & Daltry , O. ocellatus (Morice) , O. saintgironsi David, Vogel & Pauwels , and O. macrurus (discussed above). Additionally, from the species of the Oligodon cyclurus group occurring in Indochina, Oligodon arenarius sp. nov. differs externally by the dorsal coloration features (all members of the Oligodon cyclurus group except O. macrurus possess blotches, reticulation or crossbars) and by the following characters: from O. cattienensis , by the immaculate belly and a lesser ventrals count (131–144 vs. 167–178); from O. chinensis , by a much lesser ventrals count (131–144 vs. 170–206); from O. fasciolatus , by DSR 17 vs. 21–23 and by a lesser ventrals count (131–144 vs. 147–210); from O. formosanus , by DSR 17 vs. 19 and a lesser ventrals count (131–144 vs. 154–189); from O. kampucheaensis , by DSR 17 vs. 15 and a lesser ventrals count (131–144 vs. 165); from O. ocellatus , by DSR 17 vs. 19, a relatively longer tail in males (TaL/TL 0.26–0.28 vs. 0.11–0.14) and a greater subcaudals count (58–60 vs. 32–44 in males, 36–40 vs. 26–33 in females); from O. saintgironsi , by a lesser ventrals count (131–144 vs. 166–184), a lesser subcaudals count in females (36–40 vs. 53) and a relatively longer tail in males (TaL/TL 0.26–0.28 vs. 0.19–0.20).
From the members of the informal Oligodon cinereus group (sensu Smith 1943, Green et al. 2010, David et al. 2011, 2012, Neang et al. 2012), which are distributed in Indochina, China, India, Andaman and Nicobar islands, and are characterized by the unforked hemipenes, not spinose but with papillae, Oligodon arenarius sp. nov. differs by the absence of a loreal (present in all group members except O. melanozonatus Wall from India) and by the following features: from O. cinereus (Günther) , by a lesser ventrals count (131–144 vs. 155–186) and by the dorsal coloration pattern (from all subspecies having blotches, reticulations or crossbars except the uniformly colored O. c. cinereus ); from O. albocinctus (Cantor) , by a lesser ventrals count (131–144 vs. 177–207) and by the dorsal coloration pattern (light crossbars in O. albocinctus ); from O. inornatus (Boulenger) , by DSR 17 vs. 15 and a lesser ventrals count (131–144 vs. 171–174); from O. joynsoni (Smith) , by a lesser ventrals count (131–144 vs. 186–195) and the dorsal coloration pattern (blotches and reticulated crossbars in O. joynsoni ); from O. maculatus (Taylor) , by a lesser ventrals count (131–144 vs. 156 – 164) and the dorsal coloration pattern (dark blotches in O. maculatus ); from O. melanozonatus , by a lesser ventrals count (131–144 vs. 171–173) and the dorsal coloration pattern (blackedged or black crossbars in O. melanozonatus ); from O. nagao David, Nguyen, Nguyen, Jiang, Chen, Teynié & Ziegler , by a lesser ventrals count (131–144 vs. 184–193), a relatively longer tail in males (TaL/TL 0.26–0.28 vs. 0.14–0.15) and the dorsal coloration (butterfly-shaped blotches in O. nagao ); from O. splendidus (Günther) , by DSR 17 vs. 21, a single pair of both prefrontals and internasals (up to four pairs of each in O. splendidus ), a lesser ventrals count (131–144 vs. 169–193) and the dorsal coloration (large spots on dorsum in O. splendidus ); from O. woodmansoni (Sclater) , by 8(7) supralabials vs. 6, a lesser ventrals count (131–144 vs. 180–190) and pale belly coloration (dark brown with white edges in O. woodmansoni ).
The members of the informal O. dorsalis group (sensu Smith 1943) are distributed in Indochina, India, Nepal and Myanmar; the species with documented hemipenial morphology are characterized by unforked and usually spinose hemipenes. From all congeners belonging to this group, except O. lacroixi and O. catenatus discussed above, Oligodon arenarius sp. nov. differs by the dorsal coloration (longitudinal stripes or crossbars in all members of the O. dorsalis group) and an entire anal plate vs. divided, as well as by the following features of the pholidosis: from O. dorsalis (Gray & Hardwicke) , by DSR 17 vs. 15, the absence of a loreal and a lesser ventrals count (131–144 vs. 162–188); from O. eberhardti Pellegrin , by DSR 17 vs. 13, 8(7) supralabials vs. 6 and a lesser ventrals count (131–144 vs. 165–191); from O. erythrogaster Boulenger , by a lesser ventrals count (131–144 vs. 163–186); from O. hamptoni Boulenger , by DSR 17 vs. 15, 8(7) supralabials vs. 5 and a lesser ventrals count (131– 144 vs. 160–175); from O. mcdougalli Wall , by DSR 17 vs. 13 and a lesser ventrals count (131–144 vs. 199).
In the species of the informal O. torquatus group (sensu Smith 1943, Green et al. 2010) with the distribution in India, Myanmar and Thailand, hemipenes are unforked with apical papillae. Oligodon arenarius sp. nov. differs from the species of this group by the dorsal coloration features (longitudinal stripes in the O. torquatus group except O. planiceps ), an entire anal plate vs. divided and additionally by the following external features: from O. cruentatus (Günther) , by a lesser ventrals count (131–144 vs. 148–173); from O. planiceps Boulenger , by DSR 17 vs. 13, 8(7) supralabials vs. 5(4), a greater subcaudals count (36–60 vs. 22–27) and the immaculate ventrals (ventrals with paired dark blotches in O. planiceps ); from O. theobaldi (Günther) , by the absence of a loreal and a lesser ventrals count (131–144 vs. 164–180); from O. torquatus (Boulenger) , by DSR 17 vs. 15 and the absence of a loreal.
In the species of the informal O. venustus – O. taeniolatus group (sensu Smith 1943, Green et al. 2010) with distribution in India, Sri Lanka, Afghanistan, Pakistan, Nepal and Bangladesh, hemipenes are unforked and spinose. Oligodon arenarius sp. nov. differs from the species of this group by the absence of spines on the hemipenis, the dorsal coloration pattern (crossbars or longitudinal stripes or blotches in all species of the O. venustus – O. taeniolatus group), an entire anal plate vs. divided, and the following external characters: from O. affinis Günther , by the immaculate belly (with black subrectangular spots in O. affinis ); from O. arnensis (Shaw) , by a lesser ventrals count (131–144 vs. 164–202); from O. brevicauda Günther , by the presence of internasals, DSR 17 vs. 15, a lesser ventrals count (131–144 vs. 158–173) and a greater subcaudals count (36–60 vs. 25–29); from O. calamarius (Linnaeus) , by the absence of a loreal and a greater subcaudals count (36–60 vs. 20–34); from O. erythrorachis Wall , by DSR 17 vs. 15, 9(8) infralabials vs. 4 and a lesser ventrals count (131–144 vs. 154); from O. melaneus Wall , by DSR 17 vs. 15, the absence of a loreal and a lesser ventrals count (131–144 vs. 152–160); from O. sublineatus Duméril, Bibron & Duméril , by DSR 17 vs. 15 and the absence of a loreal; from O. taeniolatus (Jerdon) , by DSR 17 vs. 15, the absence of a loreal and a lesser ventrals count (131–144 vs. 158–218); from O. travancoricus Beddome , by a lesser ventrals count (131–144 vs. 154–155); from O. venustus (Jerdon) , by the immaculate belly (with large black spots in O. venustus ).
From other species of the genus, which are not clearly assigned to any specific groups of species, Oligodon arenarius sp. nov. differs by the following characters: from O. ancorus (Girard) (the Philippines), by the dorsal coloration (large dorsal spots in O. ancorus ), the absence of a loreal and a lesser ventrals count (131–144 vs. 147– 173); from O. annulifer (Boulenger) ( Malaysia: Borneo), by the dorsal coloration (crossbands in O. annulifer ), DSR 17 vs. 15 and the absence of a loreal; from O. bitorquatus Boie ( Indonesia) , by the dorsal coloration (red and yellow dots in O. bitorquatus ) and the immaculate belly (with black spots in O. bitorquatus ); from O. booliati Leong & Grismer (Peninsular Malaysia), by the dorsal coloration (crossbands in O. booliati ) and 9(8) infralabials vs. 7; from O. everetti Boulenger ( Malaysia and Indonesia), by DSR 17 vs. 15 and the absence of a loreal; from O. forbesi (Boulenger) ( Indonesia) , by the absence of a loreal, a lesser ventrals count (131–144 vs. 150–170) and the immaculate belly (ventrals with brown spots in O. forbesi ); from O. jintakunei Pauwels, Wallach, David & Chanhome ( Thailand) , by the dorsal coloration (crossbars in O. jintakunei ), DSR 17 vs. 15, an entire anal plate and a lesser ventrals count (131–144 vs. 189); from O. kheriensis Achardji & Ray ( Nepal and India), by the absence of a loreal and a lesser ventrals count (131–144 vs. 196); from O. lungshenensis Zheng & Huang ( China) , by DSR 17 vs. 15, 8(7) supralabials vs. 6, 9(8) infralabials vs. 6 and a lesser ventrals count (131–144 vs. 163–179); from O. meyerinkii (Steindachner) (the Philippines and Malaysia), by the absence of a loreal and a lesser ventrals count (131–144 vs. 154–169); from O. modestus Günther (the Philippines), by DSR 17 vs. 15, 8(7) supralabials vs. 6 and a lesser ventrals count (131–144 vs. 158–176); from O. nikhili Whitaker & Dattatri ( India) , by DSR 17 vs. 15, the dorsal coloration (longitudinal stripes in O. nikhili ) and the immaculate belly (ventrals with black spots in O. nikhili ); from O. notospilus Günther (the Philippines), by the absence of a loreal and the dorsal coloration (large rhombic yellow spots in O. notospilus ); from O. octolineatus (Schneider) ( Malaysia and Indonesia), by the dorsal coloration (longitudinal stripes in O. octolineatus ), the absence of a loreal and a lesser number of ventrals (131–144 vs. 155–197); from O. ornatus Van Denburgh ( China and Taiwan), by the dorsal coloration (crossbars in O. ornatus ), DSR 17 vs. 15 and a lesser number of ventrals (131–144 vs. 156–182); from O. perkinsi (Taylor) (the Philippines), by the absence of a loreal and a lesser number of ventrals (131–144 vs. 183–188); from O. petronellae Roux ( Indonesia) , by the dorsal coloration (dark spots in O. petronellae ), DSR 17 vs. 15 and the absence of a loreal; from O. praefrontalis Werner ( Indonesia) , by the presence of internasals, an entire anal plate vs. divided and a lesser number of ventrals (131–144 vs. 183–193); from O. propinquus Jan ( Indonesia) , by the absence of a loreal and DSR 17 vs. 15; from O. pulcherrimus Werner ( Indonesia) , by the dorsal coloration (elongate spots on dorsum in O. pulcherrimus ), the absence of a loreal and a lesser number of ventrals (131–144 vs. 152–176); from O. purpurascens (Schlegel) ( Thailand, Malaysia and Indonesia), by the absence of a loreal, DSR 17 vs. 19–21 and a lesser number of ventrals (131–144 vs. 150–210); from O. signatus (Günther) ( Indonesia and Malaysia), by the dorsal coloration (yellow spots on dorsum in O. signatus ) and the absence of a loreal; from O. trilineatus (Duméril, Bibron & Duméril) ( Indonesia) , by the dorsal coloration (longitudinal stripes in O. trilineatus ) and the absence of a loreal; from O. unicolor (Kopstein) ( Indonesia) , by the absence of a loreal and a lesser number of ventrals (131– 144 vs. 162); from O. vertebralis Günther ( Indonesia and Malaysia), by the dorsal coloration (white or yellow dorsal spots in O. vertebralis ), DSR 17 vs. 15 and the absence of a loreal; from O. waandersi (Bleeker) ( Indonesia) , by DSR 17 vs. 15 and a greater number of subcaudals (36–60 vs. 18–29); from O. wagneri David & Vogel ( Indonesia) , by DSR 17 vs. 15 and the dorsal coloration (white crossbars in O. wagneri ).





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.