
Kairona selva Huber & Carvalho, 2019
|
publication ID |
https://doi.org/10.11646/zootaxa.4546.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:D2C9F49A-9B76-40AE-9A60-CAE9B99BA547 |
|
DOI |
https://doi.org/10.5281/zenodo.5449741 |
|
persistent identifier |
https://treatment.plazi.org/id/E21587DB-FFAC-FFEC-FF11-FB884E2EFEE0 |
|
treatment provided by |
Plazi |
|
scientific name |
Kairona selva Huber & Carvalho |
| status |
sp. nov. |
Kairona selva Huber & Carvalho View in CoL sp. n.
Figs 198–212 View FIGURES 198–204 View FIGURES 205–206 View FIGURES 207–212
Gen.n. Br16-196: Eberle et al. 2018 (molecular data); Huber et al. 2018: fig. 3.
Type material. BRAZIL: ♂ holotype, UFMG (22740) ; 1♂ 1♀ paratypes, UFMG (21856, 22741); 1♂ 1♀ paratypes, CHNFUPI (2483, 2526); and 2♂ 1♀ paratypes, ZFMK ( Ar 20632), Amazonas , Tabatinga , forest at the Brazilian army base Comando de Fronteira Solimões, 8° Batalhão de Infantaria ( 4.245°S, 69.923°W), 90 m a.s.l., 2–4.xi.2016 ( B.A. Huber, L.S. Carvalho) GoogleMaps .
Other material examined. BRAZIL: 2♂ 1♀, CHNUFPI (2492, 2536, 2556) ; 2♀, UFMG (21835, 22622); and 2♀ 1 juv., ZFMK (Br16-322), all in pure ethanol, same data as types .
Etymology. The species name is derived from Portuguese selva = jungle, which is the watchword of the 8th Infantry Battalion of the Jungle of the Brazilian army (who hosted us at Tabatinga); noun in apposition.
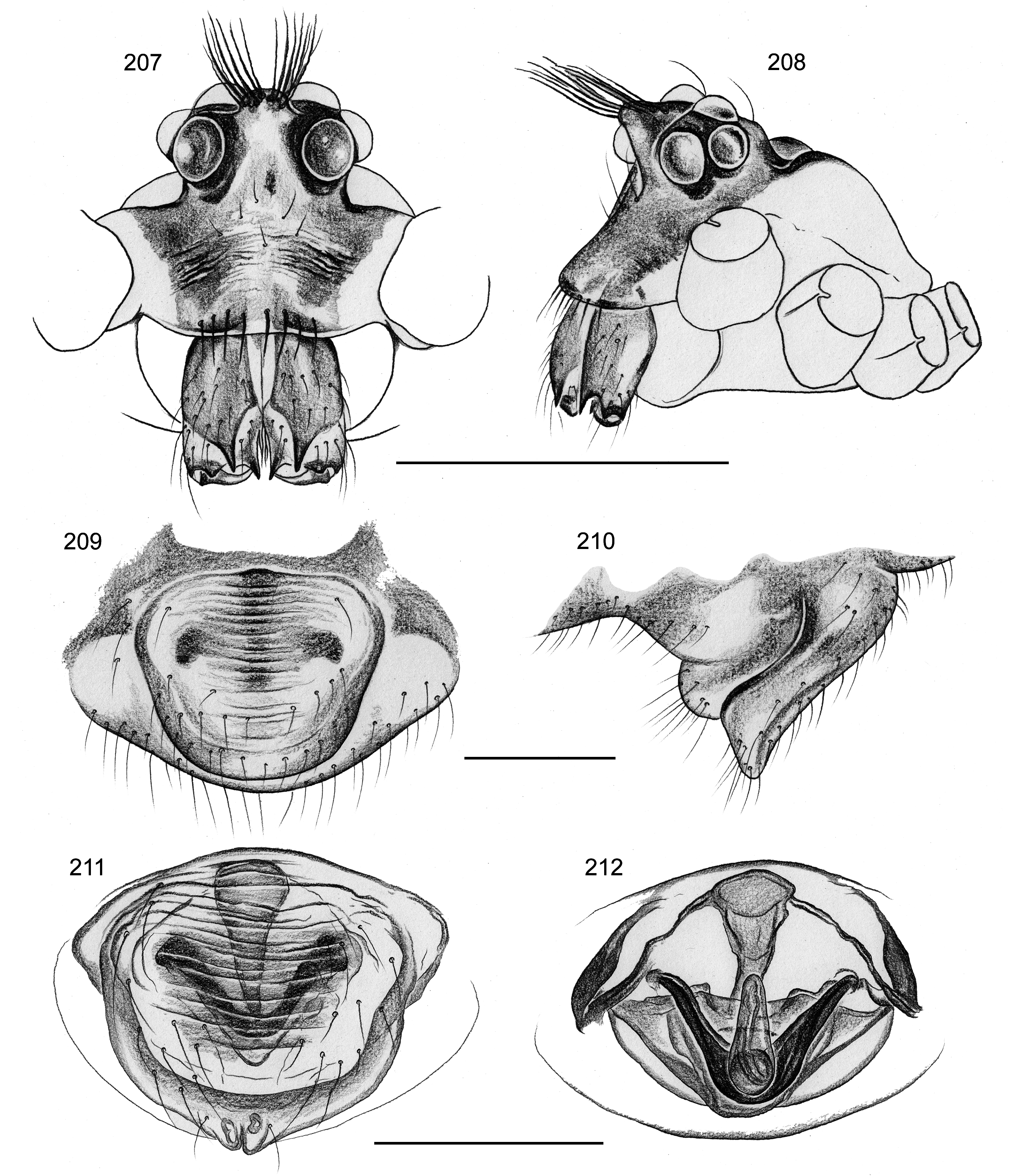
Diagnosis. Morphologically, the genus/species differs from most other known Pholcidae by shape of abdomen in dorsal view ( Figs 198–201 View FIGURES 198–204 ; wide and obtuse posteriorly, similar only in Giloloa gen. n. and in some species of Metagonia , both representing Pholcinae ); also by very distal position of metatarsal trichobothrium (on male metatarsus 1 at ~40%; see Note below); from putatively closest relatives ( Saciperere , Pisaboa , Waunana ) also by absence of AME. Males are also easily distinguished from all other known pholcids by unique brush of strong hairs on short median horn anteriorly on ocular area ( Figs 207–208 View FIGURES 207–212 ); also by distinctive shape of procursus ( Fig. 206 View FIGURES 205–206 ; sclerotized distal element with pointed processes) and by armature of chelicerae ( Figs 207–208 View FIGURES 207–212 ; pair of pointed apophyses pointing downwards). Females are also easily distinguished by internal genitalia ( Figs 204 View FIGURES 198–204 , 211–212 View FIGURES 207–212 ; distinctive V-shaped structure and putative ducts in median plane).
Description. Male ( holotype). MEASUREMENTS. Total length 1.4, carapace width 0.63. Distance PME-PME 65 µm; diameter PME 80 µm; distance PME-ALE 40 µm; AME absent. Leg 1: 17.8 (4.3 + 0.2 + 4.3 + 7.6 + 1.4), tibia 2: 2.4, tibia 3: 1.6, tibia 4: 2.0; tibia 1 L/d: 86.
COLOR (in ethanol). Carapace pale ochre-yellow, darker medially and around ocular area; clypeus also darker; sternum whitish; legs pale ochre yellow with brown rings in patella area and distally on tibiae. Abdomen gray, with three pairs of dark purple marks dorsally.
BODY. Habitus as in Figs 198–199, 201–202 View FIGURES 198–204 . Ocular area elevated, with distinctive median horn with strong hairs ( Figs 207–208 View FIGURES 207–212 ). Thoracic furrow deep. Clypeus unmodified but with row of stronger bristles. Sternum wider than long (0.44/0.32), unmodified. Abdomen distinctively angular in dorsal view.
CHELICERAE. As in Figs 207–208 View FIGURES 207–212 , with one pair of simple frontal apophyses pointing downwards.
PALPS. As in Figs 205–206 View FIGURES 205–206 ; coxa with retrolateral process, trochanter barely modified, femur with retrolateral process proximally; procursus distally strongly sclerotized, tip strongly curved towards prolateral; bulb large, with membranous and sclerotized elements.
LEGS. Without spines and curved hairs; with vertical hairs in higher than usual density on all tibiae; retrolateral trichobothrium of tibia 1 at 11%; tibia 1 without prolateral trichobothrium (present on other tibiae); tarsus 1 with>20 pseudosegments only distally fairly distinct. Metatarsal trichobothrium in very distal position (on metatarsus 1 at 42%).
Male (variation). Tibia 1 in 2 other males: 4.1, 4.7. Metatarsus 1 trichobothria in 2 other males at 37% and 39%, respectively.
Female. In general similar to male ( Fig. 200 View FIGURES 198–204 ) but without horn in ocular area and without vertical hairs on leg tibiae. Tibia 1 in 5 females: 2.9-3.2 (mean 3.1). Epigynum simple externally ( Figs 209–211 View FIGURES 207–212 ), weakly sclerotized, with pair of membranous short processes medially at posterior rim. Internal genitalia complex, with prominent Vshaped structure and putative ducts in median plane ( Figs 204 View FIGURES 198–204 , 212 View FIGURES 207–212 ), pore plates in unusual position ( Fig. 204 View FIGURES 198–204 ).
Note. Metatarsal trichobothria are in a very proximal position (i.e. at <10% of metatarsus length) in most or all pholcid species studied previously. This character appeared quite invariable in a first cladistic analysis of the family ( Huber 2000) and was thus not coded. Since then, hundreds of further species have been examined by the first author. The proximal position of the metatarsal trichobothrium appeared universal and was thus neither quantified nor included in species descriptions/redescriptions. As a result, it is not entirely clear if the present species is indeed unique for its distal position of the metatarsal trichobothrium or not. In the putatively close relatives, the trichobothria are in the usual proximal position (at 8% of metatarsus length in Pisaboa silvae Huber, 2000 ; at 5% in Saciperere catuaba sp. n.).
Distribution. Known from type locality only ( Fig. 345 View FIGURE 345 ).
Natural history. This species was found in humid forest leaf litter, on the undersides of dead leaves on the ground. Two egg sacs were weakly enveloped with silk lines and contained 12 and 19 eggs, respectively.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.