
Kerivoula krauensis, Francis & Kingston & Zubaid, 2007
|
publication ID |
https://doi.org/ 10.3161/1733-5329(2007)9[1:ANSOKC]2.0.CO;2 |
|
DOI |
https://doi.org/10.5281/zenodo.4323652 |
|
persistent identifier |
https://treatment.plazi.org/id/E079879B-C66E-FFA8-C3F9-FE0F6E493DA7 |
|
treatment provided by |
Valdenar |
|
scientific name |
Kerivoula krauensis |
| status |
sp. nov. |
Kerivoula krauensis View in CoL sp. nov.
( Figs. 1–2 View FIG View FIG , Table 1)
Holotype
Natural History Museum (formerly known as British Museum Natural History — BMNH) 99.294 (field No. CMF 911010.1), ♀ collected 10 Oct 1991. Body in ethanol, skull extracted.
Type Locality
Kuala Lompat, Krau Wildlife Reserve, Pahang, peninsular Malaysia (3°43’N, 102°10’E). Paratypes
Department of Wildlife and National Parks, Malaysia (PERHILITAN), un- catalogued, field No. TK960704.1, ♁ collected 4 July 1996; Senckenberg Museum SMF 83824 (field No. CMF920707.3) ♀ collected 7 July 1992, both from same loca- tion as holotype, body in ethanol, skulls ex- tracted.
An additional 29 males and 27 females from Krau Wildlife Reserve captured be- tween 1996 and 2004 were referred to this species in the field based on external appearance, of which most were released, but two males (TK020921.1 and TK040122.1) and one female (TK020624.2) were kept as vouchers and are in the collection at PERHILITAN.
Etymology
Named for Krau Wildlife Reserve, the locality in peninsular Malaysia where it has
in apparent colour from the first specimen are due largely to differences in lighting
been found, and ensis (Latin for ‘belonging
to’). Its proposed English name is ‘Krau
woolly bat’.
Diagnosis
A small Kerivoula , similar in size and form to K. hardwickii , but differing in dis- tinctive colouration, fur having extensive black bases with tips that range from shiny gold to light buffy-brown dorsally, and whitish-buff ventrally, and slightly smaller skull with more inflated braincase, shorter rostrum, shorter canines and more rounded upper premolars. DNA barcode differs by about 11% from all other currently recog- nized Kerivoula , a difference comparable to or greater than that among other species pairs within the genus.
Measurements
We measured the type series and a com- parative series of K. hardwickii from penin- sular Malaysia (Table 1). Field measure- ments of additional, referred specimens (29 ♁♁, 27 ♀♀) were as follows: forearm (mm), ♁♁ 0 = 29.9 (range 28.2–33.3), ♀♀ 0 = 30.6 (range 28.9–32.1); body mass (g), ♁♁ 0 = 3.0 (range 2.0–4.5), ♀♀ 0 = 3.4 (range 2.0–4.5).
Description
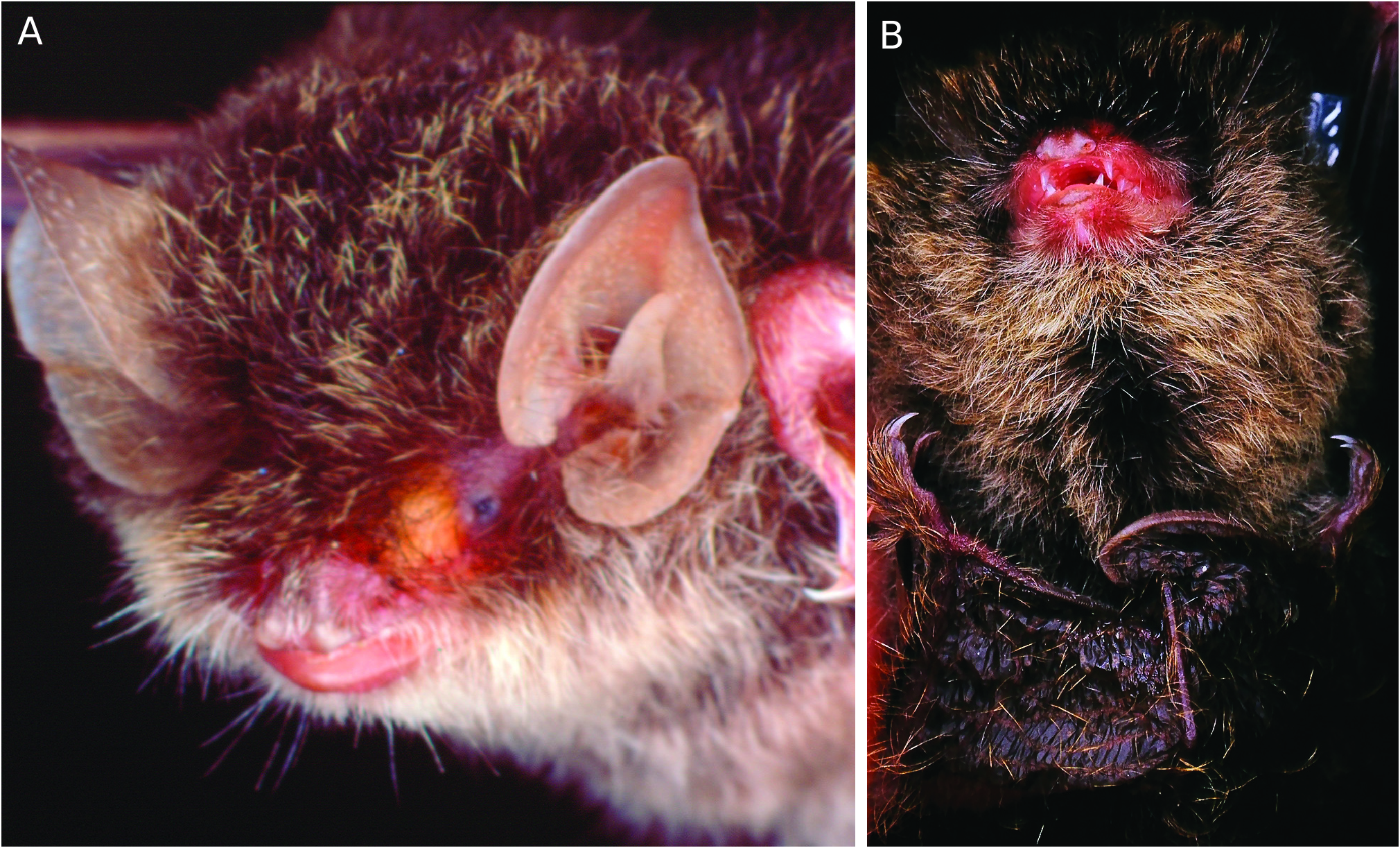
Fur on upperparts is long and woolly, dark brown for 90% of its length with shin- ing golden-brown tips ( Fig. 1 View FIG ). Fur tips on some of the referred individuals from the same locality are a duller buffy-brown, pos- sible related to age, wear, or individual var- iation. Fur on underparts has dark grey bases for about 70–80% of the length of the hairs with relatively broader greyish white to grey brown tips. Ears are moder- ately short, broadly funnel shaped with a distinct fold just posterior of the tip of the ear, the posterior flap nearly equal in width to the anterior flap such that the edges more or less meet if the ear is folded in half. The
ears are pale pinkish grey, slightly darker around the rims and paler at the base. Tra- gus is tall and pointed, curved slightly out- wards, lightly pigmented, the same general colour as the ear. The face, nose and lips are pinkish to purplish brown except for a yel- lowish gland extending about 60% of the way from the eye to the nose. The wing and tail membranes are dark brown. The wing membrane is inserted on the foot just below the base of the toes. The feet are broad, cov- ered with long golden hairs, with relatively long claws. The interfemoral membrane is sparsely covered with long, dark brown hairs with gold-tips, as on the back. The wing membranes are devoid of long hairs on the dorsal surface.
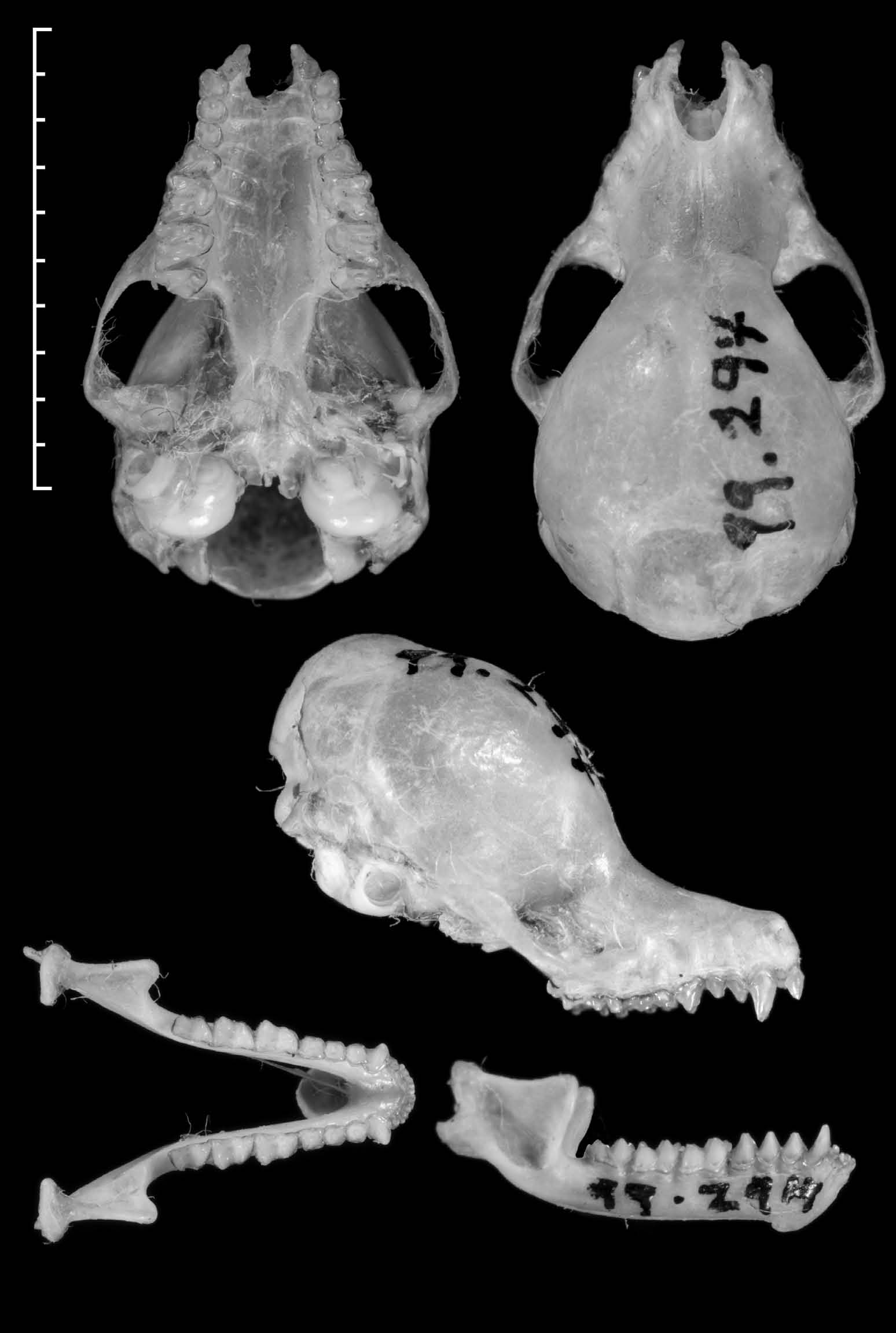
The skull shape and dentition ( Fig. 2 View FIG ) are generally similar to K. hardwickii , though slightly smaller (Table 1). The braincase is large and globular, the rostrum is relatively short with a distinct median rostral depression. The upper canine is nar- row and moderately tall, about 50% longer than the posterior premolar (P4), without any accessory cusps. The anterior incisor (I2) is conical and unicuspid, slightly less than 50% of the height of the canine; the second incisor (I3) is small and short, less than half the height of I2 but similar in crown area. The anterior premolars are both similar in size, with P2 slightly taller than P3, both reaching to about half the height of P4. In cross section, they are both rounded, slightly wider than long, of similar crown area to each other, and about half the crown area of P4 which is somewhat triangular in shape. The upper molars have well devel- oped W-shaped cusps, as is typical for the genus, the second molar (M2) distinctly wider than it is long. The lingual shelf of both M2 and M3 is broadly U-shaped with rounded edges.
In the lower jaw, the canine is relatively short, about 30% higher than the anterior lower premolar (P2) which is about 5–10% shorter than P3 and P4; the latter two are approximately equal in height to each other. Viewed from above, the two anterior premolars P2 and P3 are approximately equal in surface area and the posterior premolar P4 is about 20% larger. In profile, the lower molars M1–3 are all fairly similar in height, with the anterior cusps about 30% higher than the posterior cusps. The posterior lower molar (M3) has the talonid (posterior part) noticeably narrow compared with the trigonid (anterior part) which has a well developed lingual shelf. The teeth of the holotype are moderately worn, but the same pattern of relative sizes is apparent in TK960704-1 which has unworn teeth.
Morphological Comparison with Other Species of Kerivoula
We compared the new species with all currently recognized forms of Kerivoula from mainland Southeast Asia and the Sunda shelf, as well as two names currently considered synonyms of K. hardwickii , the most similar species from this region.
Kerivoula hardwickii is most similar in overall size, shape and morphology to the new species, but is larger (Table 1) with longer ears, different fur colour, less shortened rostrum and several differences in dentition. The type specimen of K. hardwickii , collected in Java, was described as having ‘very long, delicate, soft silky fur, grayish brown above, and brown, with a tawny tint, underneath’ ( Horsfield, 1824). Specimens that we have examined from Borneo, peninsular Malaysia, peninsular Thailand, and Lao PDR all tend to fit with this general description, although there is moderate geographic variation in fur colour on the upperparts, ranging from pale grey to light brown. DNA analyses (see below) suggest these may be a complex of species, but regardless, all specimens we have examined differ in fur colour from the new species in that the dark bases of the fur on the upperparts extend only about 1/3 of the length of the hairs, with the majority of the hair pale grey or brown.
Cranially, we compared the new species with the holotype of K. hardwickii (BMNH 79.11.29.181) as well as three specimens re- ferred to K. hardwickii from peninsular Malaysia (Table 1). The skull of the holo- type is badly damaged, but the rostrum, in- cluding both upper toothrows, is more or less intact. In K. hardwickii , the teeth are somewhat larger and heavier than those of the new species; the canine is relatively longer, about twice the length of P4; the pos- terior incisor, I3 is short, but with a notice- ably greater surface area than I2; the lingual shelf of M1 and M2 is broad with squared corners, making the whole tooth more rec- tangular than U-shaped. In the lower jaw, the anterior premolars P2–3 are both similar in height, slightly higher than P4 (as op- posed to shorter or equal), while the canine is about 50% longer. The teeth of the holo- type are moderately worn, but the same dif- ferences were apparent in the three speci- mens of K. hardwickii examined from peninsular Malaysia (as listed in Table 1). From these specimens, it was also apparent that K. hardwickii has a slightly less inflat- ed braincase than the new species, lacks any rostral depression, and has a distinct sagit- tal crest. Kris Helgen (personal comm.) ran a PCA analysis of the skull measurements for the new species against a data base he has accumulated of skull measurements of additional specimens referred to K. hard- wickii from Java, Borneo, West Sumatra and Sulawesi, confirming that K. krauensis is distinctly smaller than all of them.
Kerivoula engana Miller 1906 was de- scribed from Pulao Dua, a small island off Enggano Island in Southwest Sumatra but was allocated to K. hardwickii in Corbet and Hill (1992) . The type description indicates the fur of the upperparts was “a dark hair- brown on basal half […] with a broad buffy- gray area between this and the broccolibrown tips” ( Miller, 1906 b: 85), quite different from the new species. Furthermore, the skull of the holotype and another specimen from the same locality is similar in size to K. hardwickii (cbl 13.0–13.1) and hence distinctly larger than the new species. Whether or not K. engana should be considered conspecific with K. hardwickii goes beyond the scope of this paper, but it is clearly distinct from K. krauensis .
Kerivoula depressa Miller 1906 was described from Biapo, Carin Hills northeast of Tounghoo in Myanmar not far from the border of northern Thailand and was also allocated to K. hardwickii in Corbet and Hill (1992) . The fur was described as “lighter and yellower” than Javan specimens of K. hardwickii ( Miller, 1906 a: 65) though the specimen had long been immersed in alcohol at the time. The hairs were described as being brown on the basal half, buff and cream-buff distally, with slightly darker tips, quite different from K. krauensis . The skull of the holotype is only slightly longer than the new species (cbl 12.35) but has a braincase distinct- ly lower and more flattened than that of K. hardwickii and hence much lower than the new species with its relatively inflated, rounded braincase.
Kerivoula intermedia , also known from Krau, is similar in size to the new species ( Hill and Francis, 1984), but differs in having orange to orange brown fur with only the bases dark; relatively short ears with a smaller posterior fold; somewhat larger and more robust teeth; anterior upper incisor (I2) smaller, about 40% height of the upper canine with a well developed secondary cusp; second upper incisor (I3) rela- tively large and narrow, nearly the same height as I2; middle upper premolar (P3) more triangular in cross-section extending in a postero-lingual direction beside P4; lower canine slightly thinner and taller,
the braincase is angled away from the camera. Scale bar indicates 10 mm
more than 50% greater in height than the premolars; anterior cusps on lower molars about twice the height of the posterior cusps; talonid of M3 about the same width as the trigonid.
Kerivoula minuta Miller 1898 , which has also been caught at Krau, differs from the new species in several of the same fea- tures of fur colour and morphology as does K. intermedia , with orange fur and relative- ly tall incisors. However, it is smaller than the new species with distinctly smaller teeth, especially the premolars; P3 is some- what triangular in cross section, extending slightly lateral to P4.
Kerivoula pellucida (Waterhouse 1845) , also known from the same locality, is larger than the new species with long light brown hairs that have pale bases rather than dark bases; the ears and flight membranes are slightly translucent pale orange; the ears are much more elongate; the braincase is great- ly inflated anteriorly such that the rostrum and front of the braincase meet at a much steeper angle; the canines are narrow and elongate, nearly twice the height of the posterior upper premolar P4.
Kerivoula picta (Pallas 1767) , recorded from many parts of southeast Asia (but not from Krau), is also somewhat larger than the new species with very distinctive col- oration (orange fur, black and orange wings) and a very high domed braincase, similar to K. picta .
Kerivoula whiteheadi is known, on the mainland, only from the holotype of K. bicolor Thomas 1904 , which was col- lected from Biserat, Jalor in southern penin- sular Thailand, and was considered a sub- species of K. whiteheadi by Hill (1965). The skull of the holotype BMNH 3.2.6.91 dif- fers from the new species in having a more inflated braincase and longer, narrower ros- trum. The dentition is very distinctive, with the anterior two upper premolars elongate, about twice as long as wide, with blade-like cusps. The inner upper incisors are also very tall, about two-thirds the height of the canines, with well developed secondary cusps. The body of the type is in alcohol and the original colour has completely faded; however, the original description indicated the underparts were white, contrasting with the browner upperparts, and the tips of the wings were also white. Specimens we have seen of K. w. whiteheadi from Borneo share the same dental and cranial characters.
The remaining species of Kerivoula currently recognized from mainland Southeast Asia and the Sunda Shelf ( Simmons, 2005) include: K. lenis (FA 37–41, ccl 14.5–15.1), K. papillosa (FA 39–49, ccl 15.4–17.1), K. kachinensis (FA 41–43, ccl ≈ 15.5), and K. flora Thomas 1914 (FA 34–36, ccl 14.0–14.4). They are all substantially larger, and none has the distinctive colouration of the new species. Phoniscus atrox Miller 1905 , also in the subfamily Kerivoulinae and occurring at the same site, is similar in overall size and shape to the new species, though slightly larger, and also has dark fur and shiny golden tips; however, its fur has four bands of colour (dark bases, then mid- brown, then dark, then golden tips), the tra- gus is white with a distinct notch at the base, and the dentition is very different, particularly the canines which are enlarged with a deep groove down the outside.
Genetic Analyses
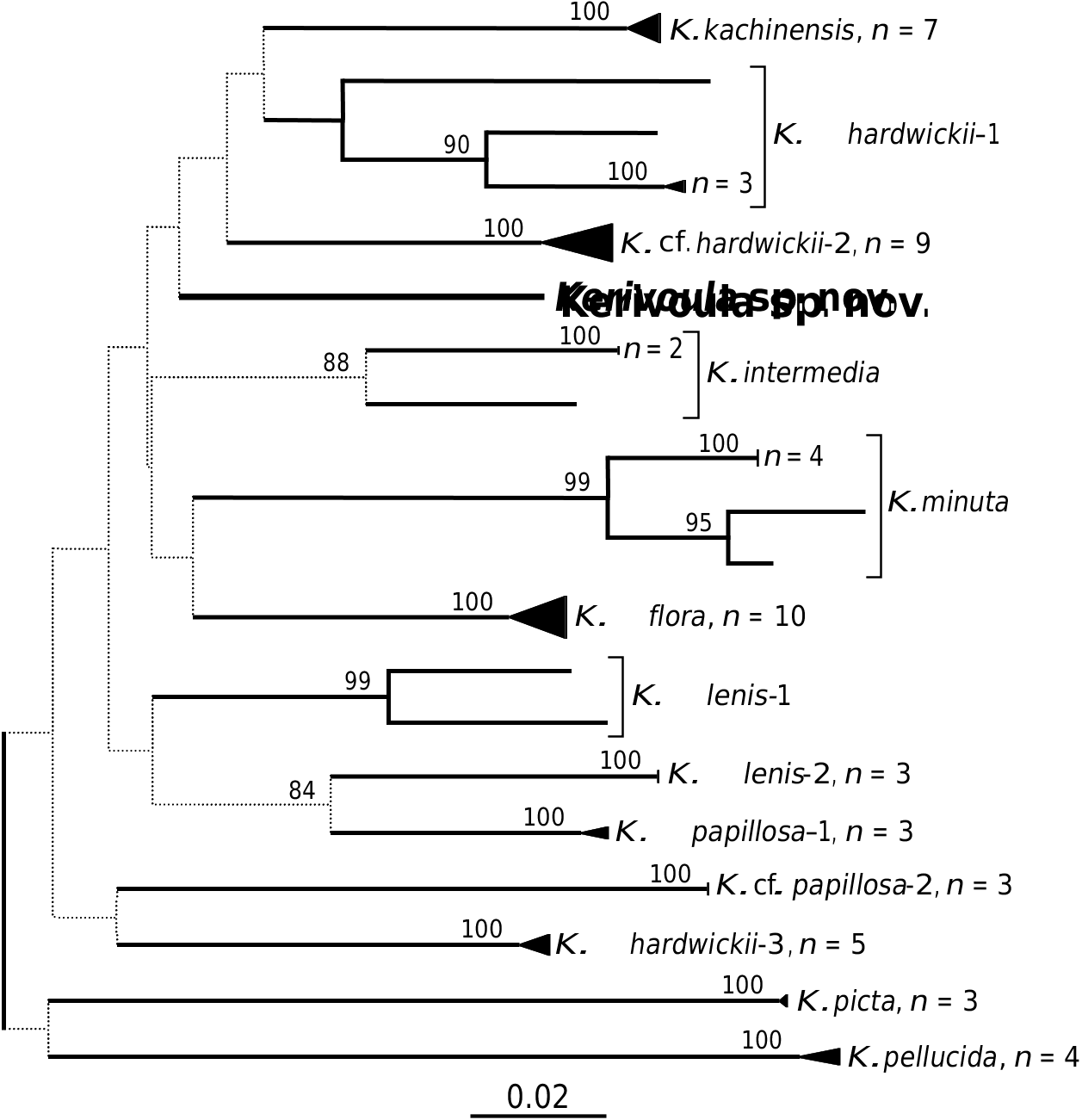
We obtained a DNA barcode for one of the paratypes (SMF 83824), as well as 63 other specimens in the genus Kerivoula rep- resenting all of the currently recognized species from mainland Southeast Asia with the exception of K. whiteheadi ( Fig. 3 View FIG ). These sequences indicate relatively low divergence within populations (represented by dark triangles or straight lines at the end of each branch) but substantial differentiation between species. The genetic distance from each species to its nearest neighbour ranged from 9.25 to 19.24%. Specimens currently
had support of <75%
referred to K. hardwickii , K. lenis and K. papillosa each included multiple clusters of DNA barcodes with divergences compa- rable to or greater than those among spe- cies, suggesting each of these may represent a complex of morphologically similar spe- cies. These clusters have been assigned ar- bitrary numbers in the graph — determina- tion of appropriate names for them will re- quire a comparative morphological analysis of the matching vouchers along with types of each named form. Specimens of K. in- termedia, K. minuta and those referred to K. cf. lenis -1 also showed moderate genetic differentiation between populations on mainland Southeast Asia and those on Borneo, but much less than differences among species.
The new species differed by 11.3% from its nearest neighbour, comparable to, or greater than the differences among other currently recognized species. The median distance from the new species to all other species of Kerivoula was 14.5%.
The genetic differentiation among species was so large for this gene that it was not
possible to determine reliably the relation- ships among species. None of the branching orders among species received bootstrap support of greater than 75%, and in most cases the support was much weaker. In or- der to obtain reliable information on phylo- genetics of Kerivoula it will be necessary to sequence additional genes (most likely nu- clear genes) that evolve more slowly.
Echolocation
Three individuals referred to this spe- cies emitted steep frequency-modulated echolocation calls characterized by very large bandwidth, high start and end fre- quencies and short duration. Echolocation calls were also of very low intensity, and sound energy was distributed fairly evenly across the frequency range. The following parameters were derived from time-expand- ed (10×) sequences (0 ± SD based upon six calls for each individual): mean call dura- tion 2.4 ± 0.8 ms, start frequency 174 ± 6 kHz, end frequency 50 ± 11 kHz. Calls were given in groups of 1–15, with a mean of 4, with an interval between calls within a group of 10–20 ms, and between groups of 39–86 ms.
Ecology
All known examples of this bat have been caught in the understorey of mature lowland rainforest. The type specimen was caught over small pools in the stream bed of a seasonal stream in relatively undisturbed forest about a kilometre from the clearing at Kuala Lompat. Other individuals were caught along forest understorey trails, showing no particular association with streams or swamps. Nothing is known of the species’ roosting ecology, although, like other Kerivoula it has not been found to roost in caves in the area. Most records of females in breeding condition (pregnant or lactating) were between February and June ( Table 2 View TABLE ), although it should be noted that trapping was rarely conducted between November and December because of the monsoon season.
| BMNH |
United Kingdom, London, The Natural History Museum [formerly British Museum (Natural History)] |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |