
Potamotrygon tigrina, De, Marcelo R., Sabaj, Mark H. & Lovejoy, Nathan R., 2011
|
publication ID |
https://doi.org/10.5281/zenodo.319945 |
|
DOI |
https://doi.org/10.5281/zenodo.5658663 |
|
persistent identifier |
https://treatment.plazi.org/id/E05B87D6-FFC3-FFA2-FF43-FC3D8EBEFAB3 |
|
treatment provided by |
Plazi |
|
scientific name |
Potamotrygon tigrina |
| status |
sp. nov. |
Potamotrygon tigrina View in CoL , n. sp.
Figures 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ; Tables 1–2 View TABLE 1
Holotype. MUSM 39978 [ex. ANSP 182452], juvenile male, 495 mm TL, Río Nanay (Amazonas drainage), large beach (left bank) at village of Pampa Chica, 4.54 km west of Iquitos, Departamento Loreto, Peru, 03°45’09” S, 073°17’00” W, collected with a bag seine by M. Sabaj, C. Pérez, A. Bullard, O. Castillo & C. DoNascimiento, 8 March 2005. Specimen dissected dorsally and ventrally by A. Bullard to remove parasites (intestine, olfactory bulbs and gills fixed); caudal stings missing when captured ( Figures 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 , 5 View FIGURE 5 , 6 View FIGURE 6 ).
Paratype. ANSP 191998, preadult female, 560 mm TL, Río Nanay, Iquitos, Departamento Loreto, Peru (specimen donated to Nathan Lovejoy by Richard Ross; specimen somewhat dry, color faded) ( Figures 4 View FIGURE 4 , 6 View FIGURE 6 ).
Diagnosis. A species of Potamotrygon diagnosed by its unique dorsal color pattern, composed of a light to dark brown or blackish background color, with a tan, bright yellow or orange irregular, highly curved and convoluted vermiculate pattern over entire disc and base of tail region, extending posteriorly to about one-half of tail length (to close to level of caudal stings). Potamotrygon tigrina , n. sp., is further distinguished from all other species of Potamotrygon , except P. schroederi , by presenting the distal half of tail, from about caudal sting origin, with a sharply contrasted pattern of five to six solid dark brown to black bars and uniformly cream colored interspaces of more or less equal width. It is further differentiated from congeners, except P. schroederi , P. orbignyi , P. h u m e ro s a Garman, 1913, P. histrix (Müller & Henle, 1834) and P. m a r i n a e Deynat, 2006, by presenting a single developed angular cartilage that is about one-fourth hyomandibula length (as opposed to two developed angulars of similar size, or a single developed but very elongated angular close to one-half hyomandibular length in other congeners). Furthermore, in P. t i g r i n a, n. sp. (and P. schroederi ), dorsal tail spines usually occur in two irregular, low, undefined rows which originate relatively far back on tail, at level more or less of disc apices, with spines relatively well spaced apart (congeners usually with a single or multiple row of dorsal tail spines, usually in well defined rows, with relatively taller tail spines that are more close packed, originating more anteriorly on dorsal tail region at tail base).
Description. Measurements, as both raw data in mm and transformed into % DW, are presented in Table 1 View TABLE 1 ; meristic features are presented in Table 2.
External morphology. Disc longer than wide, relatively oval, and widest at more or less midlength. Disc length ranges between 105 and 112.8% DW (x = 108.9% DW). Snout with oval anterior margin in dorsoventral view, and with minute but clearly visible rostral knob protruding from anterior disc. Preorbital snout length slightly more than twice interorbital distance; preorbital snout length from 26.8 to 27.1% DW (x = 27% DW). Prenasal (16 to 18.2% DW, x = 17.1% DW) and preoral (22 to 23.6% DW, x = 22.8 % DW) snout lengths relatively short, shorter than preorbital snout length. Eyes relatively small, but clearly protruding from top of head region; eyes smaller than spiracles in diameter. Spiracles closely adjacent to eyes, rhomboidal in shape and very large in live specimens, slit-like in preserved holotype. Spiracles in fresh specimens about as long as wide (length measured diagonally), longer than wide in preserved holotype. Spiracles without elevated spiracular rims or central knob posteriorly. Spiracles extend anteriorly to level of posterior two-thirds of eye length in preserved material, but in live specimens spiracles extend anteriorly beyond level of eyes. Interorbital and interspiracular distances about equal.
Mouth relatively small, its opening somewhat straight across. Mouth width about one-half distance between first gill slits, and slightly greater than internarial distance. Posterior profile of lower jaw marked with faint grooves due to preservation, but labial folds absent; outer corners of mouth also with slightly marked grooves. Nostrils anteroposteriorly elongated and slit-like, close in length to eye diameter and about one-half of internarial distance. Nasal curtain widest posteriorly, with highly fringed, medially notched posterior margin. Teeth set in quincunx, not visible externally with mouth closed in holotype. Tooth rows 42/ 46 in holotype (lower symphysial row 23), 43/ 46 in slightly larger paratype; teeth relatively small, rhomboidal, but without conspicuous cusps (holotype a juvenile male). Five buccal papillae present inside mouth. Branchial basket longer than wide, its length apparently about one-fifth DW (proper measurements could not be taken; Table 1 View TABLE 1 ). Distance between first gill slits slightly greater than distance between fifth gill slits. Gill openings with slightly undulated gill flaps; fifth gill slit smallest, and slightly more obliquely positioned.
Pelvic fins anteroposteriorly short, much wider than long (59.3 to 60% DW, x = 59.7% DW), and with undulating posterior margins. Pelvic fins mostly concealed in dorsal view, and broadest at more or less their anterior third, with broadly oval apices. Tail somewhat robust at base, its width slightly greater than interorbital distance (tail width 15.6 to 18.2% DW, x = 16.9% DW). Tail tapering posteriorly, not terminating as an elongated whip distally. Tail very flat ventrally, noticeable in cross-section. Tail with slender, ridge-like lateral tail folds originating at base of tail and extending to more or less level of caudal sting; dorsal and ventral tail fold present, ridge-like, originating posterior to caudal stings and extending caudally for a short distance. Dorsal and ventral tail folds anteriorly wider, extending posteriorly as a very shallow ridge. Cloaca to distal tail length varying greatly (108.1 to 131.2% DW, x = 119.7% DW), and greater than disc length and snout to cloaca length (87.2 to 91.2% DW, x = 89.2% DW); tail relatively intact (i.e. not broken distally) in holotype. Caudal sting (missing from holotype) much greater than interorbital distance or tail width at base, relatively elongate. Caudal sting with posteriorly directed and sharp lateral serrations, terminating in a very acute distal point, and positioned just posterior to tail mid-length.
Coloration. Dorsal surface of disc, in alcohol preserved holotype, with dark to light brown background color, becoming slightly lighter close to disc margin. Tan, yellow to orange irregular vermiculated pattern over entire disc and base of tail posteriorly to about one-half of tail length, close to level of caudal stings. Vermiculations finest in interorbital region, coarsest on posterior portion of disc, middisc region and dorsal tail base ( Figures 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 ). Isolated, well-defined ocelli lacking; however, some larger tan spots enclosed by narrower ring of dark gray to brown evident at posterior middisc and near base of tail (spots integrated into overall vermiculate pattern). In general, dorsal vermiculate pattern highly curved and convoluted. Outer disc margin with yellow to creamy white small spots (close in size to eye diameter). Outer disc spots may coalesce forming darker reticulate patterns of background color on disc margin. Dorsal surface of pelvic fins similarly with slender dark gray background covered with yellowish or creamy white spots affording coarse vermiculate pattern as spots become coalesced; spots on dorsal pelvic fins varying in size. Dorsal vermiculate pattern gradually transitions to more regular dark saddles separated by lighter, tan interspaces on basal half of tail (to origin of caudal sting). Distal half of tail, posterior to caudal sting origin, with sharply contrasted pattern of five to six solid dark bars and uniformly pale, cream colored interspaces of more or less equal width. Dark bars extend dorsally to caudal finfold margin, appear rectangular in dorsal view; extend to midline of flattened ventral surface, appearing broadly rounded in ventral view. Dark bars staggered, each one approximately aligned with pale region contralaterally on both tail and dorsal finfold. Extreme tip of tail uniformly dark. Ventral surfaces of disc and pelvic fins uniformly pale, cream colored. Flattened portion of tail to base of caudal sting with dark marks, continuous with dorsal and lateral darker bars, on both sides; these alternating darker bars do not connect ventrally. Posterior margins of disc and pelvic fins without darker color.
In live specimens, overlying color tannish bronze to golden yellow or even orange, with very dark brown to blackish background color; vermiculate pattern isolating dark brown to black spots of background color. Variation in vermiculations great, ranging from very tight, minute vermicular markings to more broad patterns highlighting greater background areas that range from circular to more elongate streaks. Black bars on distal half of tail more strongly contrasted with pale white to yellowish interspaces. Live newborn pups with more faded dorsal color, but vermiculate markings usually light orange.
CHARACTER A B precaudal vertebrae 24 26 caudal vertebrae 95 110 total vertebrae 119 136 diplospondylous vertebrae 93 105 upper tooth rows (radiograph) 42 – lower tooth rows (radiograph) 46 46 upper tooth rows (specimen) 42 42 lower tooth rows (specimen) 46 46 propterygial radials 35 34 mesopterygial radials 14 15 metapterygial radials 24 28 total pectoral radials 73 77 pelvic radials 20 22
Dermal denticles. Interspiracular region with very minute, dispersed denticles. Region posterior to interspiracular space also with scattered denticles, but these becoming more numerous and greater in size. Denticles over dorsal disc concentrated over middisc, extending posteriorly to posterior disc and base of tail region. Outer disc margin smooth, devoid of denticles. Denticles over dorsal disc vary in size, but always greater towards midline and somewhat evenly spaced. Denticles over dorsal midline low and variable in shape, some resembling asterisks, others with fewer, small radiating keels converging on crown apex. Dorsal midline denticles in various irregular rows, and more spaced apart than smaller denticles lateral to midline. Larger, more acute spines lacking on dorsal disc region. Dorsal midline denticles become larger over base of tail region, at about level of pectoral fin axils. Larger, rounded, dorsal tail denticles present at base of tail in paratype. Slightly taller dorsal tail spines occur relatively posteriorly on tail, just anterior to or at level of disc apices, and extend caudally in irregular rows ( Figures 5 View FIGURE 5 C, 6). In paratype, a single more defined row of dorsal tail spines present well posterior to level of pectoral axil. In holotype, dorsal tail spines slightly lower than in paratype, not forming well-defined row, and occurring in two irregular rows in which more developed spines alternate, not occurring side-by-side ( Figure 6 View FIGURE 6 ). From midtail posteriorly to caudal stings, dorsal tail spines occur in a single more defined row in holotype. Dorsal tail spines relatively well separated, not bunched together. Lateral tail denticles, much lower than dorsal tail midline spines, present on posterior tail near caudal stings, and sometimes posterior to stings, especially in paratype.
Skeletal features. Neurocranium elongate, longer than twice greatest width at nasal capsules and postorbital processes ( Figure 5 View FIGURE 5 A). Nasal capsules broadly rounded, with slender internasal septum. Preorbital processes slightly wider than postorbital processes. Precerebral and frontoparietal fontenellae very elongate, about fourthfifths of neurocranial length, leaving very anteroposteriorly short otic roof posteriorly at parietal fossa. Supraorbital process broadly triangular, just anterior to postorbital process. Preorbital processes slender. Antorbital cartilage very slender and elongate, wider anteriorly, extending posteriorly to level of angular cartilage. Meckel`s cartilage stout with well developed dorsally projecting lateral process, palatoquadrate slender. Hyomandibulae relatively slender and more or less straight, anteriorly only slightly curved toward midline and faintly concave. Anterior angular cartilage well developed, about one-third to one-fourth length of hyomandibula ( Figure 5 View FIGURE 5 B), with concave anterior margin; posterior angular absent (if present, highly reduced and uncalcified). Cervicothoracic synarcual elongate, its greatest width just under greatest width of neurocranium, with 29 spinal nerve foramina on each side. Thoracolumbar synarcual very slender, not strongly calcified. Individual vertebral centra occurring slightly posterior to level of missing caudal sting origin; a slightly calcified notochordal extension (cartilaginous rod) present distally ( Figure 6 View FIGURE 6 ). Transition from mono- to diplospondyly occurs at second centra posterior to pelvic girdle. Propterygium widest posteriorly, more stout than meso- and metapterygium. Metapterygium more broadly arched than propterygium, with four posterior segments. Anterior segment of propterygium elongate, about equal in length to nasal aperture. Mesopterygium anteriorly elongate and slender, slightly convex externally. Pectoral radial elements sometimes fused at base, especially those at posterior propterygium and anterior metapterygium. Pectoral radials slender close to pectoral basals, slightly widening and shortening toward middisc, and slender again distally, bifurcating at 9th or 10th segment. Pelvic girdle with concave anterior margins lateral to prepelvic process (broken in holotype). Lateral prepelvic processes low. Iliac processes extending caudally slightly beyond triangular ischial processes. Three to four obturator foramina present. Basipterygium relatively wide and short, but longer than posterior metapterygial segments. First enlarged pelvic radial element articulating with iliac region of puboischiadic bar. Pelvic radials articulating with posterior basipterygium splayed distally.
Remarks. Potamotrygon tigrina has been misidentified as Potamotrygon menchacai Achenbach, 1967 in the aquarium literature (e.g. Ross & Schäfer, 2000; numerous websites), even though P. menchacai was described from Río Colastiné, a tributary of Río Paraná, near Santa Fe, Argentina. This identification was based on the lightly banded distal tail thought to be present in the holotype of P. m e n c h a c a i ( Ross & Schäfer, 2000, p. 44; but see Achenbach, 1967, figure 1, p. 6), even though clearly distinct from the broad, alternating darker and lighter bars in the distal tail of P. tigrina . However, a recent systematic revision of Potamotrygon falkneri Castex & Maciel, 1963 , based on large series of specimens including material similar to the holotype of P. menchacai in color and dimensions, confirmed that P. menchacai is a junior synonym of this highly variable species ( Silva & Carvalho, 2011), as previously reported by Rosa (1985) and Carvalho et al. (2003). The holotype of Potamotrygon tigrina is very close in size to the holotype of P. m e n c h a c a i (now lost; but see Achenbach, 1967), which facilitates comparisons (note that the former is a juvenile male whereas the latter is an adult male); furthermore, Silva & Carvalho (op. cit.) thoroughly characterized P. falkneri . Potamotrygon tigrina differs significantly from this species in coloration (dorsal color composed of numerous spots or very slender vermicular markings, never forming the specific patterns of P. tigrina , and tail never similarly banded in P. fa l kn e r i), dorsal tail spines ( Figures 3 View FIGURE 3 C, 4B) (spines relatively taller, more greatly developed in P. falkneri ), configuration of suborbital loop formed by the infraorbital lateral line canal (relatively less expanded anteriorly in P. falkneri , no acute angles anteriorly; see below), single developed angular cartilage ( Figure 5 View FIGURE 5 A) (two developed angulars of roughly equal size in P. falkneri ), size at sexual maturity (smaller in P. falkneri ), among other features ( Silva & Carvalho, 2011).
Variation in dorsal color among specimens of Potamotrygon tigrina , judging from images available in the aquarium literature and other specimens we have examined, appears to be significant, but not as great as in other species, such as P. falkneri , P. m o t o ro, and P. orbignyi . Furthermore, specimens depicted always have a conspicuous pattern that allows for their identification as P. t i g r i n a, principally regarding the specific arrangement and color (strong yellow and orange in live specimens) of their intricate dorsal vermiculated design and broad banding of the posterior tail region. Some larger specimens of P. t i g r i n a have a slightly finer vermiculate pattern dorsally, but most specimens present a broader vermiculate pattern. Elevated chromatic variation is to be expected given what we know of other highly patterned species of Potamotrygon , such as in a new species being described from the mid to upper Rio Tapajós.
Potamotrygon tigrina has been imported by commercial ornamental fish dealers for at least 13 years, and is considered a difficult species to keep in captivity ( Ross & Schäfer, 2000). Allegedly, it is capable of reaching sizes of up to 1 m in disc width (which would make it one of the largest species of Potamotrygon ). The impact on this species stemming from this trade is unknown. Much work on its basic biology, including its reproductive parameters and population dynamics, is needed in order to properly evaluate if conservation measures should be enacted. Presently, all that can be determined concerning its reproduction is that the holotype (495 mm TL, 271 mm DL, and 258 mm DW) is a juvenile male with barely noticeable claspers ( Figure 3 View FIGURE 3 D); we suspect that sexual maturity in males begins at about 400 mm DW based on other specimens seen but not preserved.
Geographic distribution. The holotype is from Río Nanay, a moderate blackwater tributary of Río Amazonas, at Pampa Chica, near Iquitos, Peru ( Figure 7 View FIGURE 7 ). Pampa Chica is a large beach of fine sand along the left bank of a hairpin curve in river, approximately 11 river km upstream from the confluence with Río Amazonas ( Figure 8 View FIGURE 8 ). The specimen was collected in a bag seine pulled near shore shortly after sunset. The aquarium literature has many photographs of specimens of this species, but usually without precise locality information. These specimens were probably sold in the Iquitos ornamental fish market, which relies heavily on stingray material from the Río Nanay and Río Tigre, another affluent of Río Amazonas farther west than Río Nanay. We suspect, however, that P. t i g r i n a is slightly more widespread in the upper Río Amazonas (according to one aquarium importer, it also occurs in the Río Putumayo as well, but this is not confirmed).
Etymology. The specific epithet tigrina derives from the Latin tigris, in reference to its conspicuous dorsal disc color and vertically striped color pattern on distal tail region. Gender feminine.
Proposed common name. Tiger ray (as in the aquarium literature).
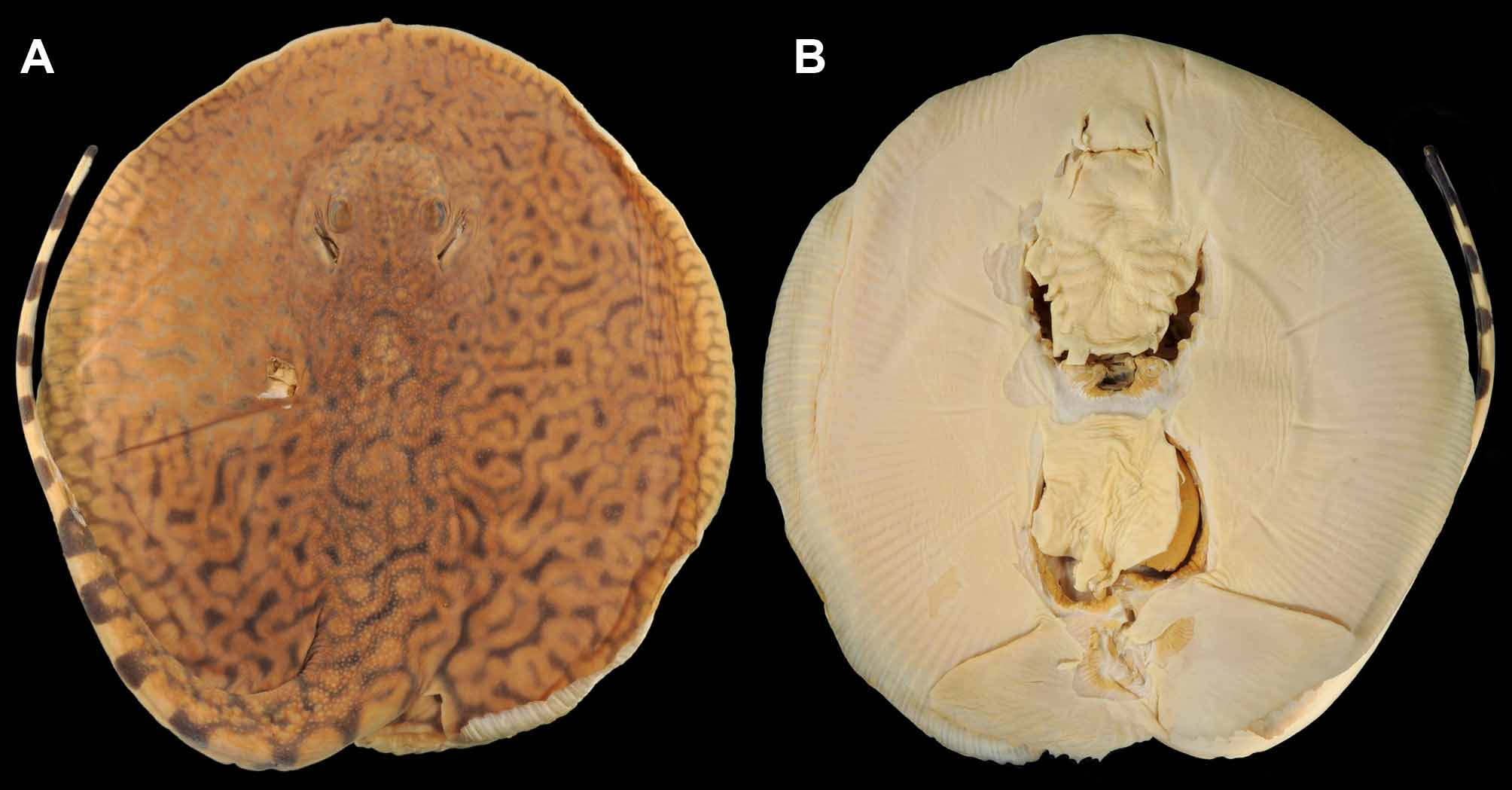

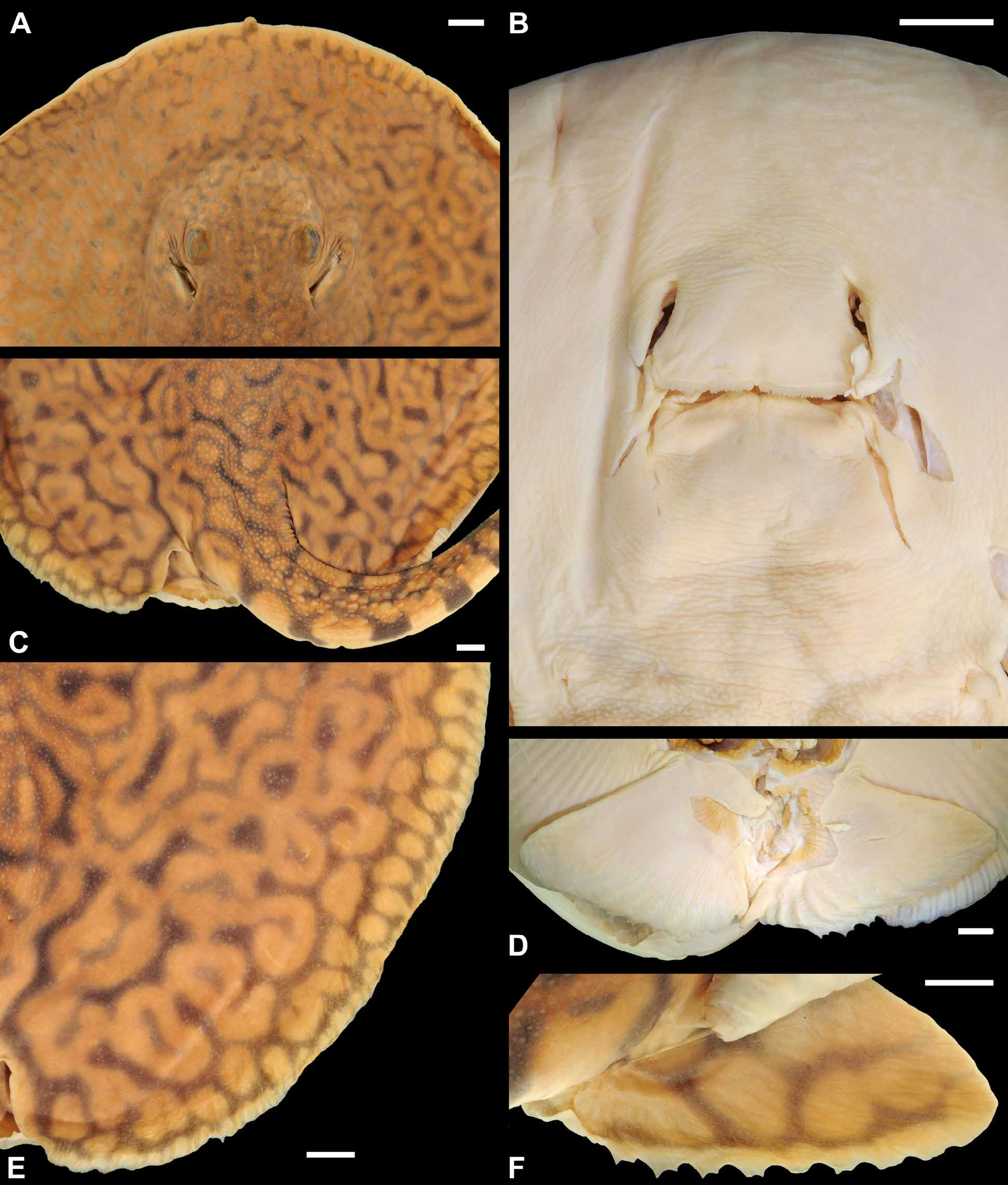
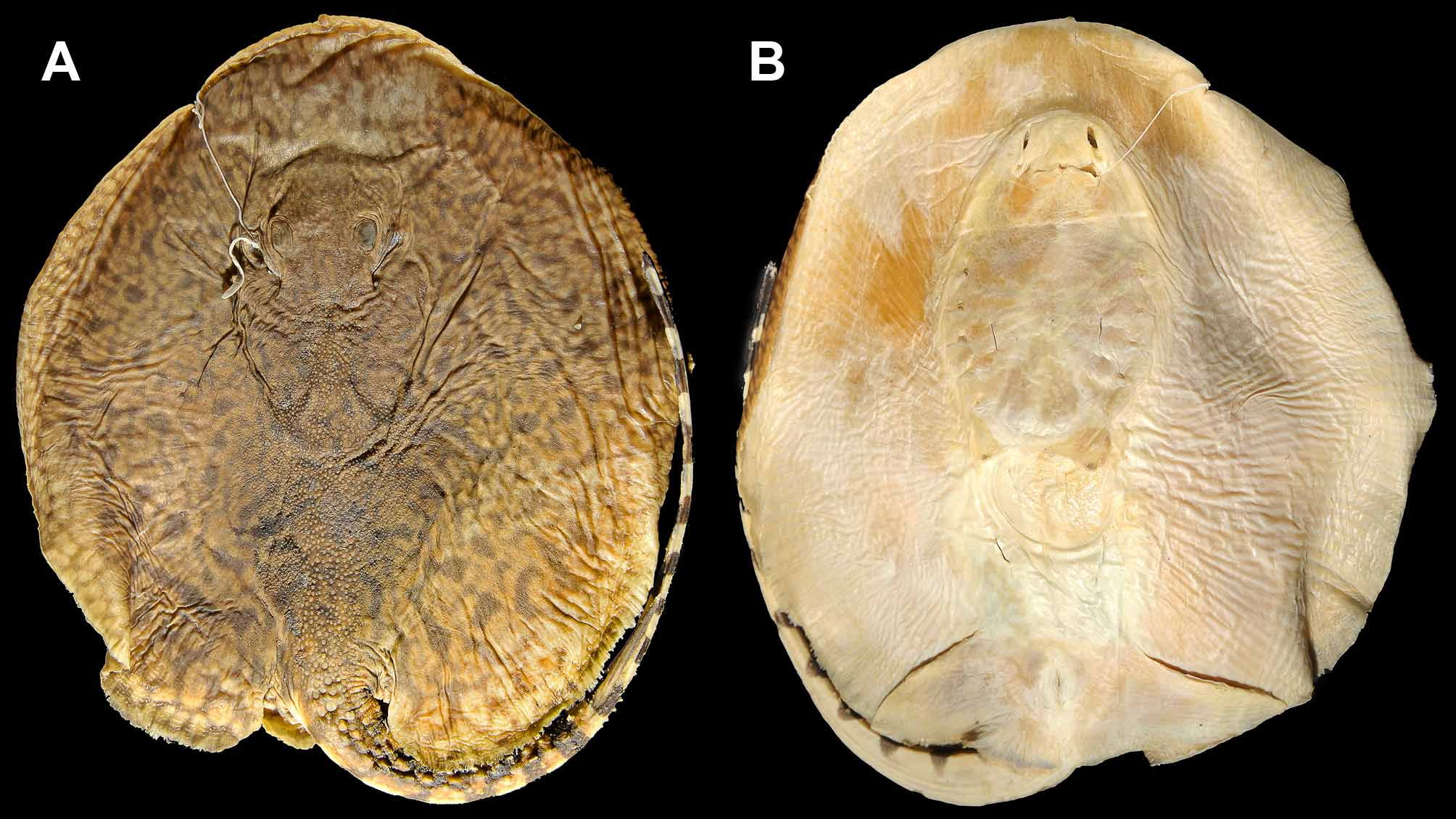




TABLE 1. Measurements of specimens of Potamotrygon tigrina, n. sp. A: MUSM 39978 (holotype), juvenile male. B: ANSP 191988 (paratype), preadult female. x: mean. * branchial basket removed in holotype; caudal stings lacking in specimen when captured.
| PARAMETER | A | B | x |
|---|---|---|---|
| mm % DW | mm % DW | mm % DW | |
| total length (TL) | 495.0 – | 560.0 – | 527.5 – |
| disc length (DL) | 271.0 105.0 | 282.0 112.8 | 276.5 108.9 |
| disc width (DW) | 258.0 100.0 | 250.0 100.0 | 254.0 100.0 |
| Interorbital distance | 34.0 13.2 | 22.0 8.8 | 28.0 11.0 |
| interspiracular distance | 43.0 16.7 | 34.0 13.6 | 38.5 15.1 |
| eye length | 22.0 8.5 | 12.0 4.8 | 17.0 6.7 |
| spiracle length | 30.0 11.6 | 21.0 8.4 | 25.5 10.0 |
| preorbital length | 70.0 27.1 | 67.0 26.8 | 68.5 27.0 |
| prenasal length | 47.0 18.2 | 40.0 16.0 | 43.5 17.1 |
| preoral length | 61.0 23.6 | 55.0 22.0 | 58.0 22.8 |
| internarial distance | 28.0 10.9 | 20.0 8.0 | 24.0 9.4 |
| mouth width distance between 1st gill slits distance between 5th gill slits branchial basket length | 33.0 12.8 * – * – * – | 24.0 9.6 56.0 22.4 40.0 16.0 47.0 18.8 | 28.5 11.2 – – – – – – |
| pelvic fin anterior margin length | 65.0 25.2 | 65.0 26.0 | 65.0 25.6 |
| pelvic fins width clasper external length clasper internal length | 153.0 59.3 6.0 2.3 34.0 13.2 | 150.0 60.0 – – – – | 151.5 59.7 – – – – |
| distance between cloaca and tail tip | 279.0 108.1 | 328.0 131.2 | 303.5 119.7 |
| tail width | 47.0 18.2 | 39.0 15.6 | 43.0 16.9 |
| snout to cloaca distance | 225.0 87.2 | 228.0 91.2 | 226.5 89.2 |
| pectoral to posterior pelvic length distance from cloaca to sting origin sting length sting width | 39.0 15.1 * – * – * – | 29.0 11.6 144.0 57.6 4.9 2.0 0.4 1.2 | 34.0 13.4 – – – – – – |
| ANSP |
Academy of Natural Sciences of Philadelphia |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Chondrichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |