
Anaspides richardsoni, Ahyong, 2016
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zlac083 |
|
DOI |
https://doi.org/10.5281/zenodo.7814230 |
|
persistent identifier |
https://treatment.plazi.org/id/D96287D1-4B67-3204-D44C-1517FB0C1612 |
|
treatment provided by |
Plazi |
|
scientific name |
Anaspides richardsoni |
| status |
|
The following descriptions explicitly refer to a detailed examination of the right Thp1 of two adult female individuals of A. richardsoni . Comparative surveys of the posterior thoracopods have been made in several specimens of A. richardsoni , Anaspides tasmaniae ( Thomson, 1893) and Anaspides clarkei Ahyong, 2015 (see Grams & Richter, 2021).
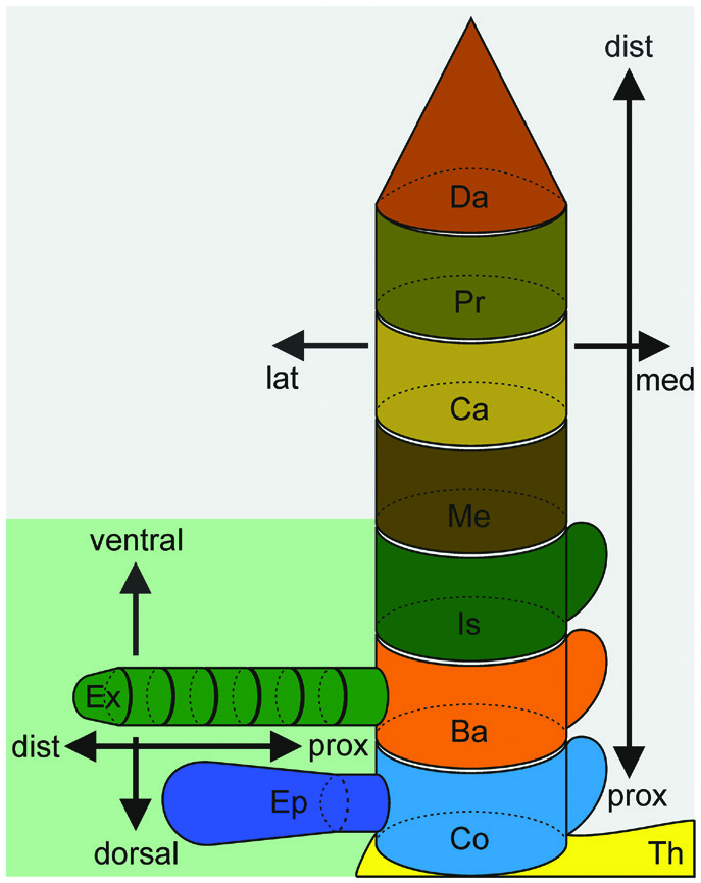
Cuticle and skeletal structures of thoracopods in Anaspides
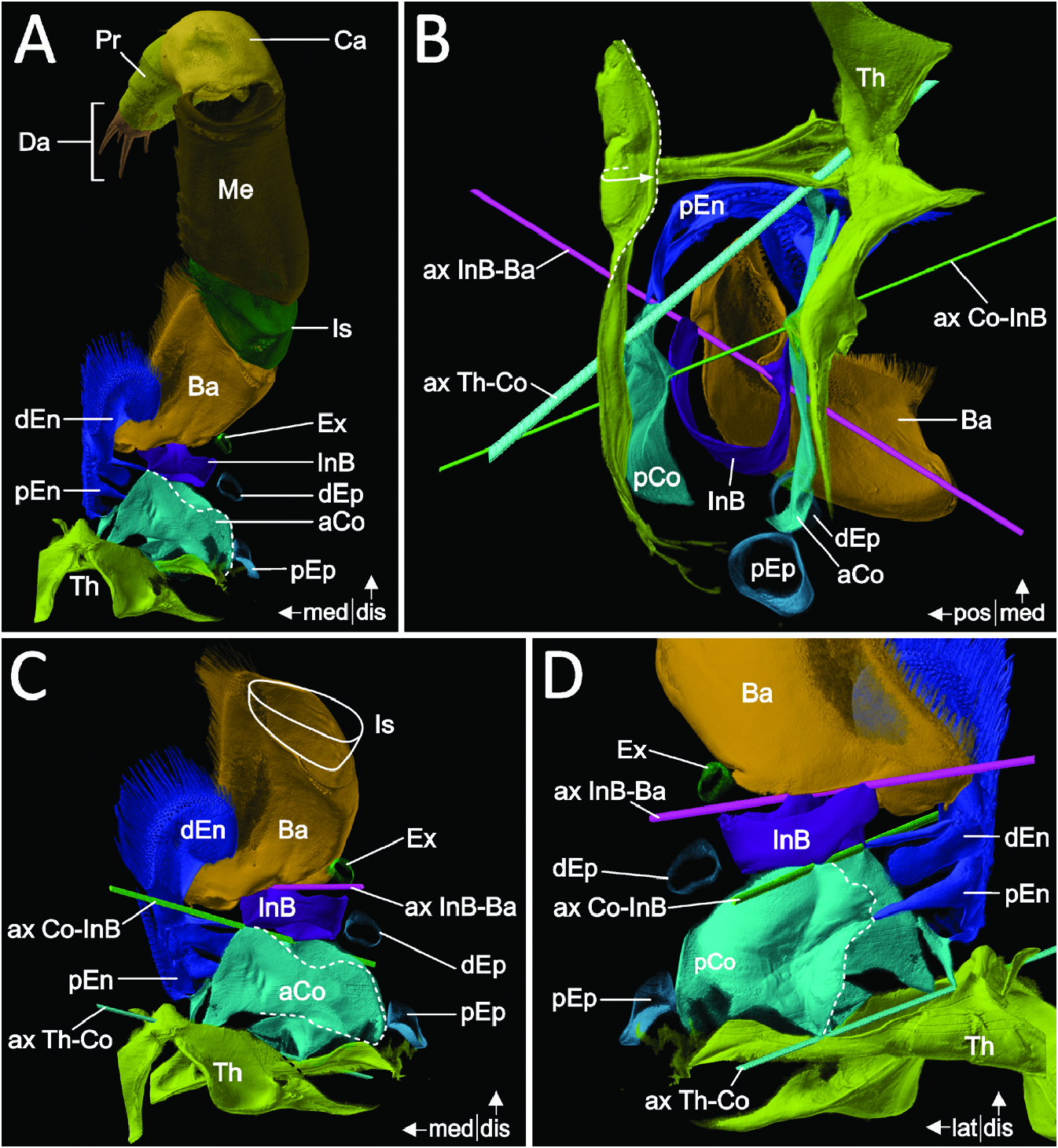
The first pair of thoracopods (Thp1) in A. richardsoni ( Fig. 2A View Figure 2 ) shows some gnathal modifications, while the following (‘posterior’) thoracopods conform to typical ‘pereiopods’. The following description refers to both types of legs, if not specified otherwise (for a detailed depiction of posterior thoracopods, see Grams & Richter, 2021).
The thorax connects and articulates with the coxa as an almost square cuticular ring. Anterolaterally, the cuticle of the thorax appears of somewhat lesser robustness ( Fig. 2B View Figure 2 ). The thorax in the posterior thoracomeres (Thp2–8, respectively) continues into a lateral plate with similar properties, whereas in the first thoracomere the thorax continues into the posterior end of a notably cuticulized carapace-like fold (a short lateral and ventrally oriented overlap of the thoracic cuticle) that reaches anteriorly up to the rostrum. Medially, the thoracic cuticle of the left and right body half is coalesced to a sternum ( Fig. 2B View Figure 2 ) that forms a shared roof-like apodeme posteromedially by being curved dorsally and anteriorly (see partly dashed curved arrow and dashed outline, Fig. 2B View Figure 2 ).
The thoracopodal protopod comprises a coxa, an intrabasis and a basis. The coxa is mainly embodied by two notably robust components: an anterior plate (aCo in Figs 2A–C View Figure 2 , 3B View Figure 3 ) and a posterior plate with a distinct invagination (pCo in Figs 2B, D View Figure 2 , 3A, C View Figure 3 ). The cuticle of the coxa is apparently less robust and more membranous in the medial to posteromedial and posterolateral to lateral areas ( Fig. 2B View Figure 2 ), but in Thp1 the medial to posteromedial area is largely covered by the robust plates and sclerites of the endites (see next paragraph). In the posterior thoracopods, the anterior coxa plate connects and articulates laterally and anteromedially with the thorax, whereas in Thp1 it is indirectly connected medially to the thorax via a sclerite of the proximal endite. The lateral articulation present in the posterior thoracopods is missing in Thp1, and the transition between the thorax and the anterior coxa appears smooth and ambiguous, without a clear interpodomere membrane. The posterior invagination of the coxa in the posterior thoracopods extends proximally into an anteriorly directed noselike apodeme. No such apodeme is present in Thp1 ( Fig. 2B View Figure 2 ). Instead, the invagination forms a posterior condyle at the proximal coxa margin that articulates with the thorax ( Fig. 2B, D View Figure 2 ). The joint constellation between thorax and coxa in the posterior thoracopods results in a lateromedial articulation axis allowing promotion and remotion. In Thp1, this axis (ax Th-Co in Fig. 2B–D View Figure 2 ) is (in comparison) rotated by almost 45°, resulting in a combined motion of promotion with abduction and remotion with adduction, respectively. On its distal margin, the coxa forms a bicondylar articulation with the intrabasis ( Fig. 2B View Figure 2 ), with an anterior condyle at the anterior coxa plate (the condyle being more strongly invaginated in Thp1; Fig. 2A–C View Figure 2 ) and a posterior condyle at the posterior coxa invagination ( Fig. 2B, D View Figure 2 ).
In females, an endite attaches medially on the coxa of thoracopods 5–7. It is lobate, moderately cuticulized and carries long, distally and slightly anteriorly directed setae. These endites are missing or at least much less prominent on the other ‘pereiopods’ and in males in general. However, on Thp1 two endites are present (in both sexes), together with a somewhat complex construction of plates and sclerites: the proximal endite appears leaf-like (pEn in Fig. 3B, C View Figure 3 ), whereas the distal endite is more or less tetrahedral (dEn in Fig. 3B, C View Figure 3 ). Both these endites have a distinct proximal plate (or stem), from which they arise (St* and St** in Fig. 3C View Figure 3 ). The plate of the proximal endite connects and articulates posterolaterally with the posterior coxa plate ( Fig. 2D View Figure 2 ; left white arrow in Fig. 3C View Figure 3 ), medially with the distal endite (right white arrow in Fig. 3C View Figure 3 ) and anteromedially, via a sclerite, with both the anterior coxa plate and the thoracic sternum ( Fig. 2C, D View Figure 2 , 3B View Figure 3 ). The plate of the distal endite reaches posterolaterally into the membranous area of the coxa (without connecting to the posterior coxa plate; Fig. 2D View Figure 2 ; St** in Fig. 3C View Figure 3 ), in addition to forming anterolaterally a shared condyle with both the anterior coxa plate and the intrabasis ( Fig. 2D View Figure 2 ; white arrow in Fig. 3B View Figure 3 ). The distal edge of the proximal endite and the distal triangular surface of the distal endite are both setose.
Laterally attached to the coxa are two leaf-like epipods, with the second epipod originating distally to the first, while being positioned anteriorly to it (pEp and dEp in Fig. 3 View Figure 3 ). In the posterior thoracopods, the epipods originate close to each other, whereas they are positioned far more apart proximodistally in Thp1 (pEp and dEp in Fig. 2A–D View Figure 2 ). Each epipod has a stem, which is clearly distinct from the distal portion. Although the epipods are only weakly cuticulized overall, the stem appears to be more strongly cuticulized than the rest. Anteriorly, the stem of the distal/anterior epipod is articulated with the coxa via a rhombus-shaped sclerite, which appears to be absent in Thp1.
The intrabasis is a robust half-ring covering the anterior, lateral and posterior wall of the leg (InB in Fig. 2B View Figure 2 ), leaving the medial side between the coxa and basis mostly membranous. A bicondylar articulation between the intrabasis and the basis (ax InB-Ba in Fig. 2B–D View Figure 2 ) is formed by an inwardly folded anterolateral edge and a posteromedial articulation point.
The basis (Ba in Fig. 2A–D View Figure 2 ) takes up about twothirds (almost three-quarters in Thp1) of the total length of the protopod. A bicondylar articulation with the endopod (i.e. the ischium) is formed by robust anterolateral and posteromedial articulation points. However, in Thp1, the posteromedial articulation point is apparently absent ( Fig. 4A, B View Figure 4 ). Here, strongly cuticulized edges are present, but without being in (at least not permanent) contact, possibly resulting in some type of ‘sliding-edge’ articulation. The proximal margin of the basis comprises a robust half-ring covering the anterolateral, anterior, medial and posteromedial edges. The rest of the basis also appears to be strongly cuticulized, although less robust compared wth the proximal margin. Both the intrabasis and basis of Thp1 generally appear to be of higher robustness than in the posterior thoracopods, with the anterior wall of the basis being prominently concave, more or less fitting the form of the distal coxal endite and the anteriorly adjacent maxilla, while the narrow medial wall is densely setose.
Within the lateral membrane between the basis and the intrabasis of Thp1–7, an exopod attaches laterally. The exopod of Thp 2–6 comprises a prominent peduncle and a long, feathered flagellum, whereas in Thp1 and Thp7 it comprises only a short stem, followed by a longish lamellar portion (Ex in Fig. 3A View Figure 3 ). In Thp2–6, the exopod articulates with the basis via an anterior hinge and a posterior rhombusshaped sclerite. Both these articulation structures are missing in Thp1.
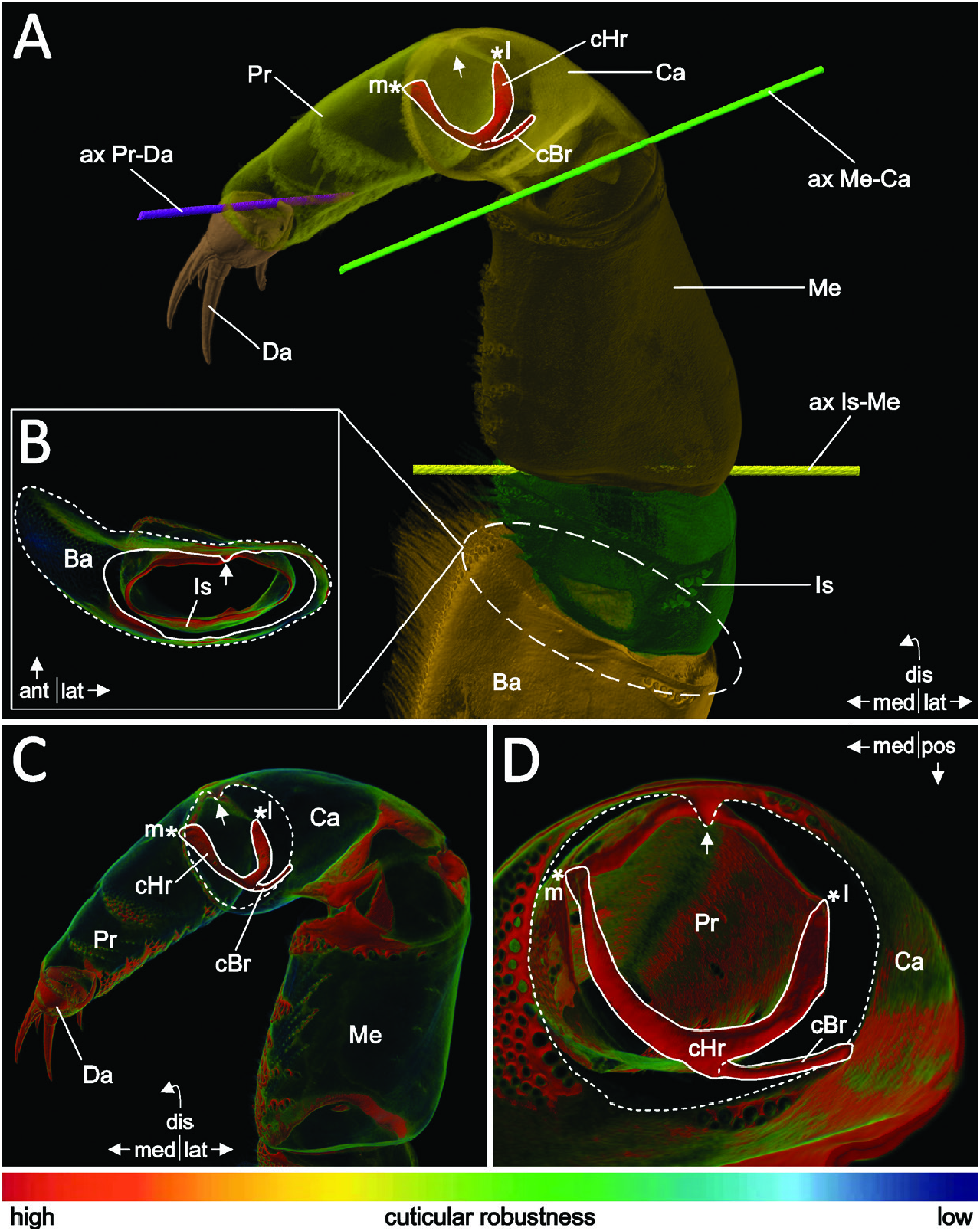
The endopod consists of the ischium, merus, carpus, propodus and dactylus and, for the most part, accords between Thp1 ( Fig. 4A–D View Figure 4 ) and the posterior thoracopods. The ischium (Is in Figs 2A View Figure 2 , 4A View Figure 4 ) measures less than half the length of the basis. A bicondylar articulation with the merus is formed by an anteromedial and a posterolateral articulation point (ax Is-Me in Fig. 4A View Figure 4 ).
The merus (Me in Figs 2A View Figure 2 , 4A View Figure 4 ) somewhat exceeds the length of the basis. A bicondylar knee-like articulation with the carpus is formed by anteromedial and posterolateral articulation points, respectively (ax Me-Ca in Fig. 4A View Figure 4 ).
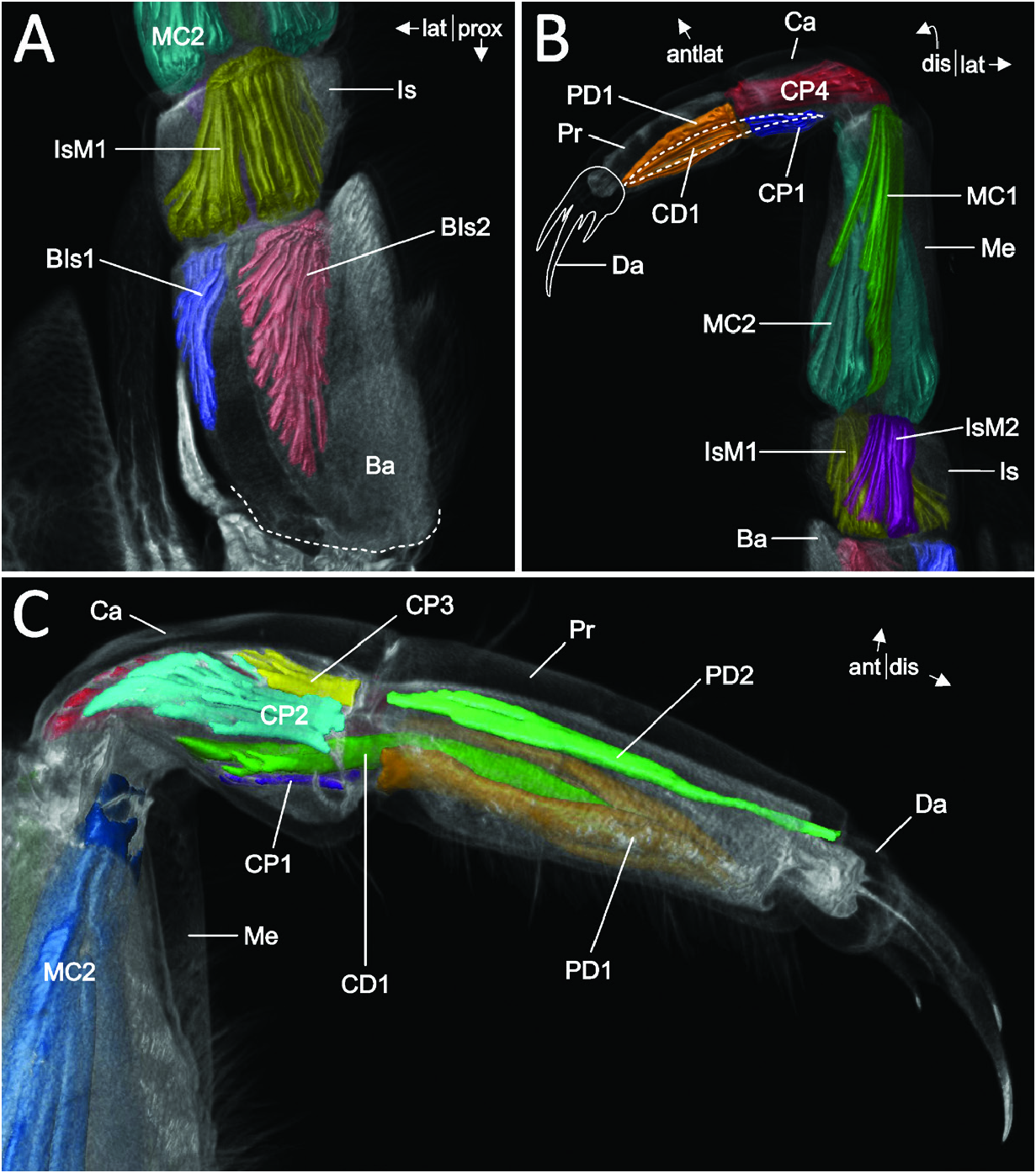
The carpus (Ca in Figs 2A View Figure 2 , 4A View Figure 4 ) measures about half the length of the basis. Posteriorly on its proximal margin, a broad sclerite reaches distally into the propodus, serving as an attachment site for muscle MC2. The carpus articulates distally with the propodus via an anterior condyle (white arrow in Fig. 4A, C, D View Figure 4 ) and a complex posterior construction: a posterolateral sclerite (cBr in Fig. 4A, C, D View Figure 4 ) connects the distal margin of the carpus to a cuticular half-ring (cHr in Fig. 4A, C, D View Figure 4 ) that is positioned posteriorly at and articulating medially and laterally with the proximal margin of the propodus (m* and l* in Fig. 4A, C, D View Figure 4 ). Two sclerites (not depicted) arise slightly proximal to the medial and lateral joints, functioning as attachment sites for muscles CP2 and CP4.
The propodus (Pr in Fig. 4A, C, D View Figure 4 ) measures about three-quarters of the length of the basis. A bicondylar articulation with the dactylus is formed by slightly posteromedial and posterolateral articulation points (ax Pr-Da in Fig. 4A View Figure 4 ).
The dactylus (Da in Fig. 4A, C View Figure 4 ) is short and of robust cuticle, ending in a lateromedial row of four (six in Thp1) strong, cuspidate setae of alternating length ( Figs 1A View Figure 1 , 3A View Figure 3 ), giving it a clawed or almost hand-like appearance. Posteriorly, on its proximal margin, a longish sclerite (not depicted) reaches into the distal third of the propodus, functioning as attachment sites for muscles CD1 and PD1.
Musculature of Thp 1 in Anaspides
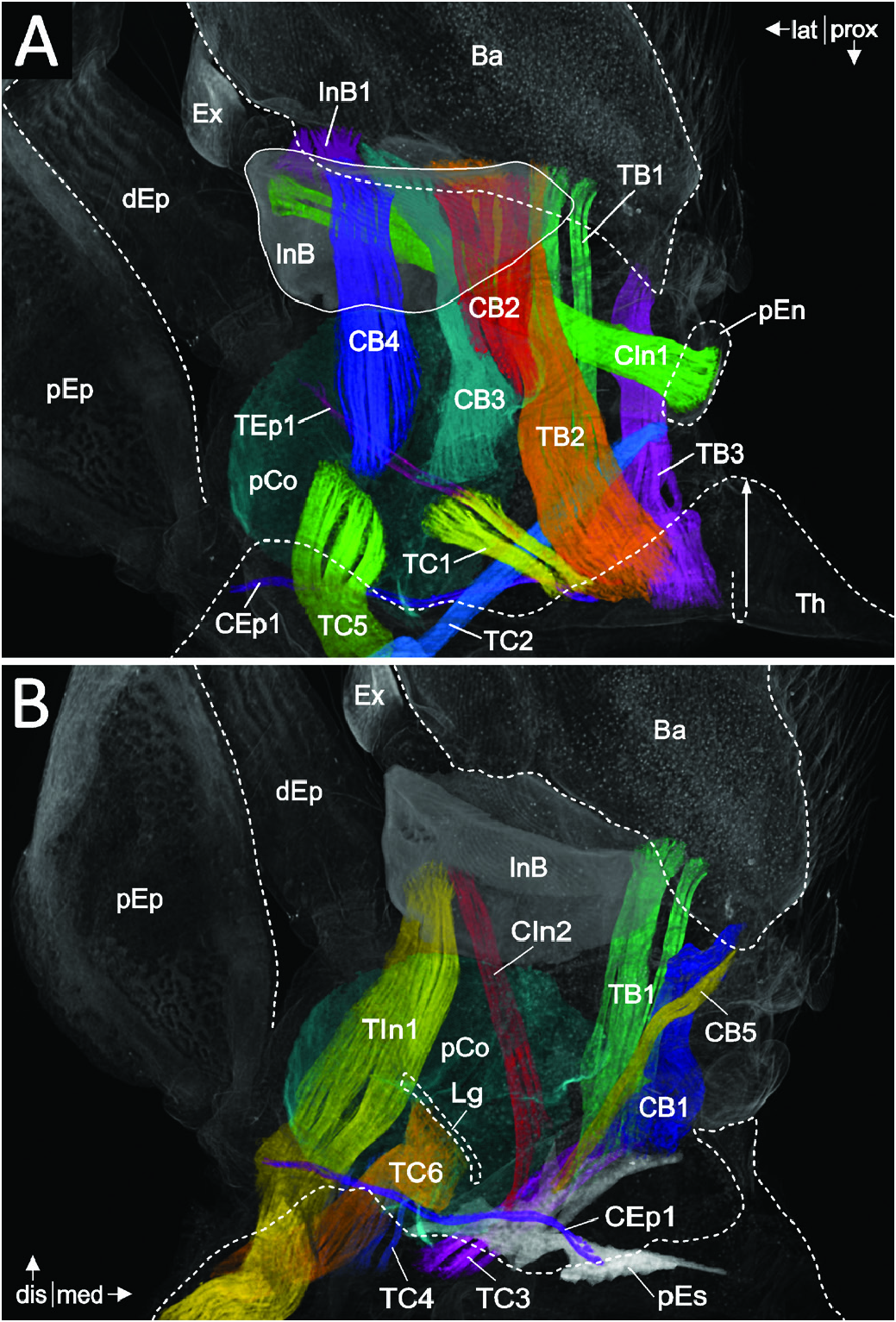
In the right Thp1, a total of 33 extrinsic and intrinsic muscles were identified ( Figs 5 View Figure 5 , 6 View Figure 6 ; for a detailed description, see Table 2 View Table 2 ).
A total of 13 extrinsic muscles connect the thorax with the leg (i.e. with the protopod; Fig. 5A, B View Figure 5 ). Six of these muscles (TC1–6) insert on the coxa, with two of them more specifically on the posterior invagination (TC1 and TC5) and one (TC6) on a ligament that connects the posterior invagination of the coxa with the slight depression of the anterior coxa wall. A further two of the 13 extrinsic muscles insert on the proximal coxal endite (TC2) and the distal epipod (TEp1), respectively. The remaining four muscles insert on the intrabasis (TIn1) and the basis (TB1–3), respectively.
Of the 20 intrinsic leg muscles, ten originate in the protopod ( Figs 5A, B View Figure 5 , 6A View Figure 6 ). Seven of these originate from the coxa (CIn1, CIn2 and CB1–CB5), with two (CIn1 and CIn2) inserting on the intrabasis and five (CB1–CB5) on the basis. One of these intrinsic muscles (CIn1) connects the proximal coxal endite with the intrabasis. Only one muscle (InB1) directly connects the intrabasis with the basis. Another two muscles originate in the basis and insert on the proximal margin of the ischium (BIs1 and BIs2). No muscles are extrinsically or intrinsically associated with the exopod.
Intrinsic to the endopod, 11 muscles are present ( Fig. 6A–C View Figure 6 ). Two of these muscles connect the ischium with the merus (IsM1 and IsM2; Fig. 6A, B View Figure 6 ) and two connect the merus with the carpus (MC1 and MC2; Fig. 6B, C View Figure 6 ). Five muscles originate in the carpus, with four of them inserting on the proximal margin of the propodus (CP1–4) and one muscle (CD1) reaching onto the proximal margin of the dactylus ( Fig. 6B, C View Figure 6 ). A further two muscles connect the propodus with the dactylus (PD1 and PD2; Fig. 6C View Figure 6 ).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |