
Polyipnus laruei, Vourey, Elodie, Dupoux, Cyndie & Harold, Antony Scott, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4263.3.8 |
|
publication LSID |
lsid:zoobank.org:pub:53BD898F-9C8F-46C9-9A02-573F3EAF41C5 |
|
DOI |
https://doi.org/10.5281/zenodo.6028485 |
|
persistent identifier |
https://treatment.plazi.org/id/D67A87E6-897E-FFB2-7A93-CFD42B3221BF |
|
treatment provided by |
Plazi |
|
scientific name |
Polyipnus laruei |
| status |
sp. nov. |
Polyipnus laruei View in CoL new species
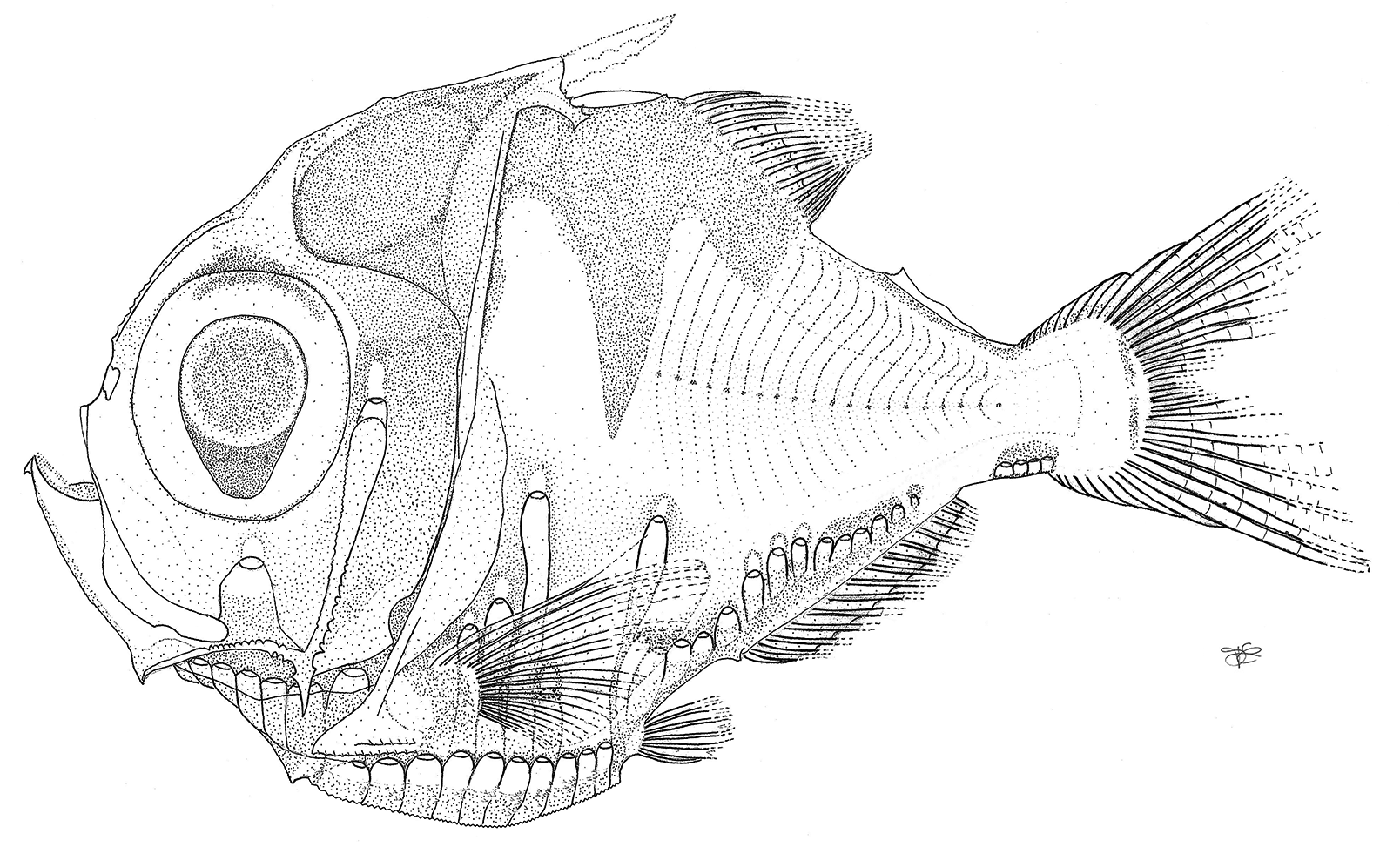
(English common name: Bigeye Hatchetfish; French name: Poisson hache à gros yeux) Fig. 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 ; Tables 1–3
Holotype. MNHN –IC– 2014–2817 ( National Museum of Natural History, France); 45.6 mm SL, 57.0 mm TL , July 30, 2014, off Reef Tombo (S 22°33.885’ latitude, E 166°25.370’ longitude), New Caledonia, western South Pacific (the only specimen of Polyipnus laruei sp. nov. was found off the southwest coast of New Caledonia, about 29 km from the nearest coastline and around 2.5 km from the barrier reef. Fishermen found the specimen dead, although in excellent condition, floating on the surface. There was no visible evidence of the cause of death). GoogleMaps
Diagnosis. A member of the Polyipnus spinosus species group with the following unique characters: presence of a single OVB photophore, as compared to other Polyipnus in which there are three. VAV, ACA and ACB photophores form a nearly continuous series in the form of a shallow, sigmoidal curve along the ventral margin of the body from the pelvic-fin base to near the insertion of the posteriormost anal-fin ray, compared with other species of the group in which there is a noticeable space between the VAV and ACA photophores (except P. fraseri in which the ACA photophores are absent but there is a gap between the VAV and ACB photophores); ACA photophores separate from each other, not united in a common organ.
The combination of the following features further serve to distinguish the new species: eye relatively large (eye diameter 22.4 % SL), presence of spines on photophore scales of PV, VAV, ACB, and ACC clusters; presence of bifurcated posttemporal spines with prominent serration; 6 ACB photophores; gill rakers 25 on left side, and 26 on right side; one long and broad lateral pigment bar extending below the lateral midline.
Description. Meristic and morphometric character values and the configuration of the photophores are given in Tables 1, 2 and 3. Body size moderate for a species of Polyipnus at 45.6 mm SL, based on the single known specimen. Dorsal-fin rays 11; anal-fin rays 15; pectoral rays 15; pelvic-fin rays 6; gill rakers 8 + 18 = 26, on right side and 8 + 17 = 25, on left side. Total vertebrae 32. Body profile anterior of dorsal- and pelvic-fin origins deep and broadly elliptical; posteriorly, body profile to caudal peduncle approximating an isosceles triangle, tapering at about 45° angle to caudal peduncle. Body very compressed, almost as deep as long. Caudal peduncle slightly elongate and rectangular, as in P. limatulus . Fin rays segmented; dorsal fin with single unbranched ray anteriorly, posteriormost ray divided to base; anal fin with three unbranched rays anteriorly; pectoral fin extending posteriorly beyond pelvic-fin base as far as third VAV photophore, and with single unbranched ray laterally; caudal fin with 10 dorsal and 9 ventral segmented rays, 9 + 8 branched rays, 10 + 8 procurrent fin rays. Adipose fin present, with long base, occupying about 61 % of dorsum between posterior dorsal-fin ray and first dorsal procurrent caudal-fin ray.
Eye large, diameter 22.4 % SL (59.0 % HL); pupils elliptical with pronounced aphakic space located ventrally to lens. Ventral margin of dentary with two keels: ventrolateral keel smooth and ventral keel with 18 serrae. Dentary teeth minute and conical to recurved anteriorly, conical on posterior portion, and differentiated as follows: four rows anteriorly, near symphysis, with number of rows decreasing posteriorly, first to three rows, then two and ultimately in a single series. Premaxillary teeth minute and conical with up to four rows anteriorly, near symphysis, with the longest row containing 28 teeth. Maxillary teeth minute, conical and uniserial with 32 teeth. Vomerine teeth in two anterolateral patches, each comprising a short, longitudinal row of four teeth. Palatines each bearing a small patch of four conical to slightly recurved teeth anteriorly. Vomerine and palatine teeth similar in size and shape, and slightly larger than adjacent premaxillary teeth.
Character Abbreviation Holotype
MNHN–IC– 2014–2817
Left side Right side
Dorsal-fin rays D 11 11 Anal-fin rays A 15 15 Pectoral-fin rays P 15 15 Pelvic-fin rays V 6 6 Caudal-fin rays C Upper Lower Segmented fin rays 10 9 Branched fin rays 9 8 Procurrent fin rays 10 8
Left side Right side
Gill-rakers GR 25 26 Lower gill-rakers LGR 8 8 Upper gill-rakers UGR 17 18 Total vertebrae 32
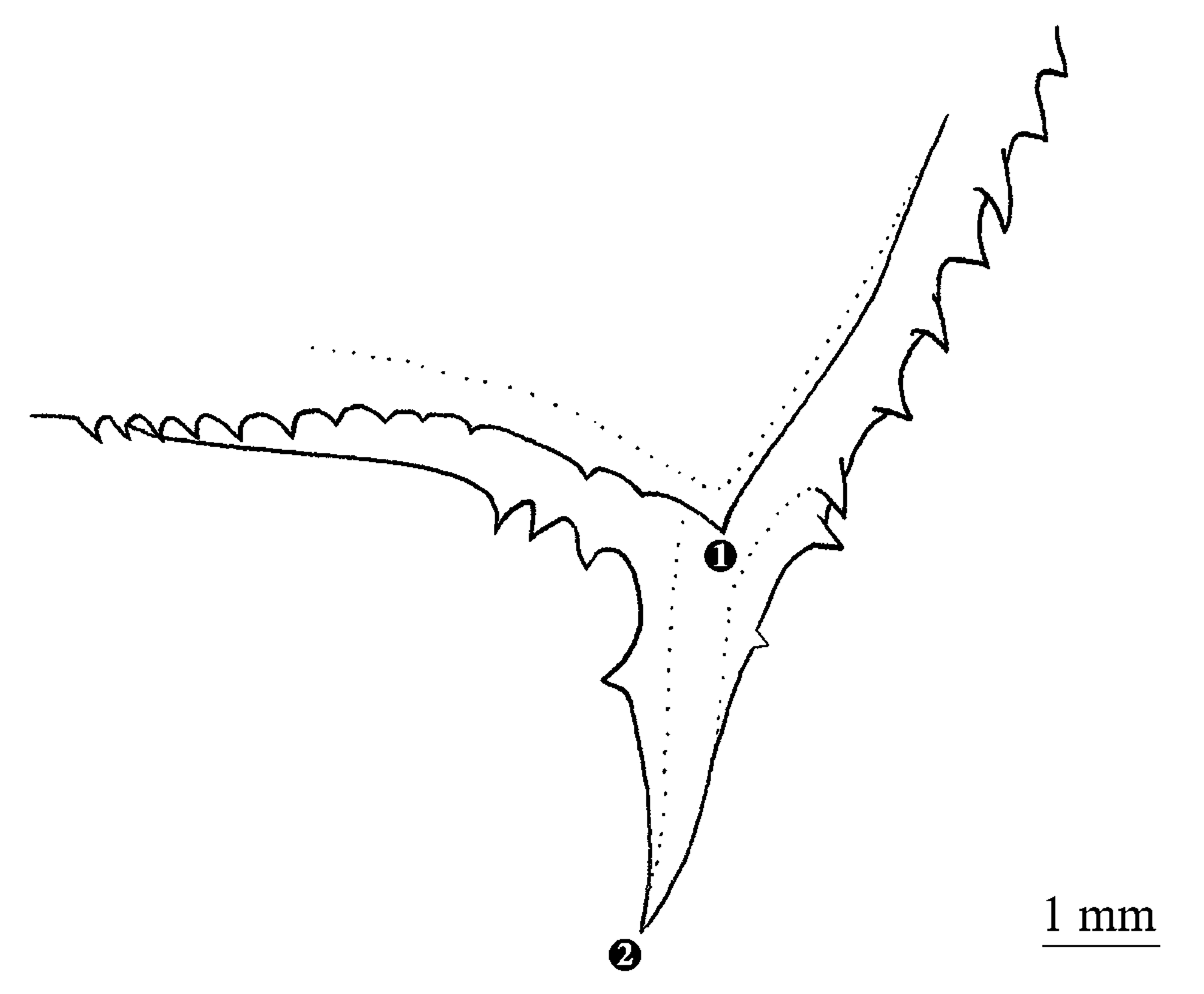
Posterior preopercular spine absent. Dorsal limb of posttemporal smooth; two prominent posttemporal spines present: dorsal spine posterodorsally directed and longest, approaching but not reaching dorsal fin origin, smooth dorsally and with distinct serration on basal two thirds of ventral margin (i.e., unarmed on upper third near apex) and basal spine a little more than half the length of dorsal spine, curving towards anterior, with serrate dorsal margin. Two preopercular spines ( Figure 3 View FIGURE 3 ): ventral spine long, about half orbit length, directed and curved anteroventrally, one serration on each side; dorsal spine short (not longer than one preopercular serration), directed laterally, without serration; posterior propercular spine absent. Vertical and anteroventral ridges of preoperculum serrate.
Dorsal spinous process present but reduced and indistinct with a bony extension of the first pterygiophore of the dorsal fin and with two approximately parallel dorsal ridges developed posteriorly as a small conical spine. Anal-fin pterygyiophore spine absent; small knob-like cartilage on each side of head of the pterygiophore.
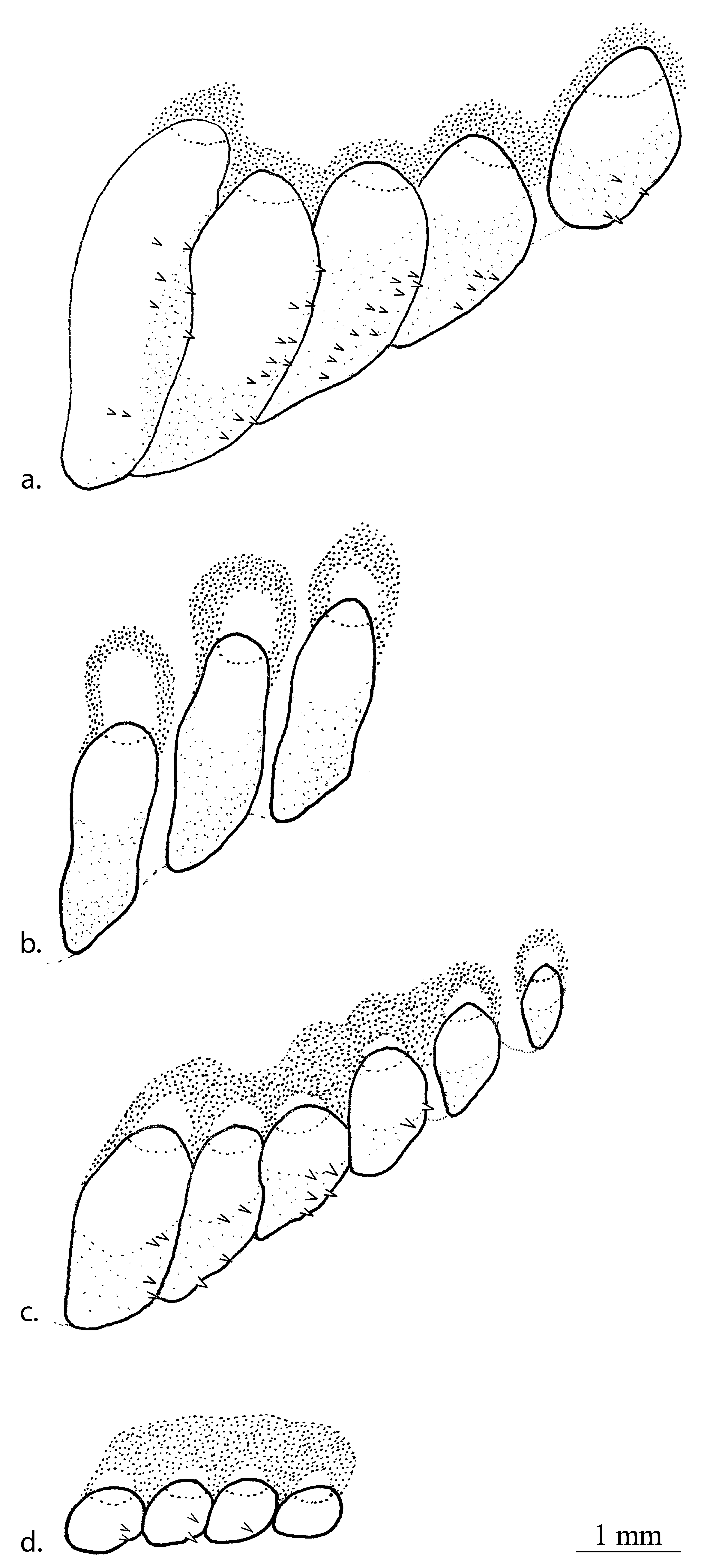
IP (5) + 1, usually (6) in other Polyipnus species; BR (6); PV (10) each ventral margin of photophores scales with numerous minute spines; ORB 1; OP 3; OVA (2) + 1, stepped dorsally from anterior to posterior, second photophore very slightly above first and the third clearly elevated to the level of ACC photophores, with each OVA photophore associated with sequential myotomes; OVB 1, with only the anterior photophore present in comparison with three OVB, as occurs in other congenerics, and separate of the third OVA by one myotome; L 1, at level slightly below third OVA; VAV, ACA and ACB phot ophores form a nearly continuous series in the form of a shallow, sigmoidal curve along the ventral margin of the body from the pelvic fin based to near the insertion of the posteriormost anal-fin ray. VAV (5), follow arc of ventral body margin, first elevated above second in cluster and fifth elevated above fourth at almost same level of first ( Figure 4 View FIGURE 4 a); ACA 1 + 1 + 1, separate, not united in a common organ, increasing in elevation from anterior to posterior. First ACA dorsally to second and third anal-fin rays, second ACA dorsally to fifth anal-fin ray, third ACA dorsally to sixth anal-fin ray ( Figure 4 View FIGURE 4 b); ACB (4) + 1 + 1, with a dorsal step between third and fourth photophore, a narrow gap between fourth and fifth, and fifth and sixth, neither fifth nor sixth photophore fully developed ( Figure 4 View FIGURE 4 c); ACC (4), compact, lacking gaps between individual photophores ( Figure 4 View FIGURE 4 d). ACB and ACC separated by broad gap equal in length to about five ACC photophores. Ventral margins of PV, VAV, ACB and ACC clusters bearing numerous spines ( Figure 4 View FIGURE 4 ); ACA photophore scales smooth, lacking spines, and in a posteriorly ascending row ( Figure 4 View FIGURE 4 ).
% SL % HL ...Continued on next page Terminology Photophores location Polyipnus laruei sp. nov.
Holotype MNHN–IC– 2014–2817
In this In Schultz Total Configuration
publication (1961)
Color in preservative. Dark brown pigment covering dorsal and dorsolateral surfaces of head and extending posteriorly as a saddle terminating near base of caudal fin. Ventral margin of dark dorsal pigment bearing undulations characteristic of most species of Polyipnus : lateral pigment bar broad-based dorsally, extending ventrally, beyond lateral midline, to a tapered point; elongated predorsal inflection present. Silvery guanine pigment covering lateral surfaces of body, and extending dorsally on either side of lateral dark pigment bar. Numerous isolated melanophores delineating myosepta and lateral midline, and on the caudal peduncle, dorsal- and caudal-fin rays. Lightly pigmented or unpigmented patch area encircled by dark chromatophores located on dorsum between the adipose fin and dorsal procurrent caudal-fin rays.
Etymology. Polyipnus laruei is named after William Larue who collected the holotype.
Remarks. The new species Polyipnus laruei is distinguished from all other species of Polyipnus by the unique presence of only one OVB photophore (the anterior photophore of the group of three that are typically present in congeners).We also report an unusual lightly pigmented patch of tissue encircled by dark chromatophores on the dorsum between the adipose fin and dorsal procurrent caudal-fin rays. This tissue is at least superficially similar to caudal glands in other stomiiforms, such as Gonostoma ( Herring & Morin 1978; Copeland 1991), although such structures are not known to occur in the Sternoptychidae .
Species of Polyipnus have been placed in four species groups: the P. asteroides , P. meteori , P. omphus and P. spinosus species groups (Harold 1994). Polyipnus laruei is considered a member of the P. spinosus species group, based on the following combination of characters: multiple posttemporal spines (two in this case) and presence of spines on the PV photophore scales. The posttemporal spines on both sides of the holotype were damaged after it was collected, however, a photograph taken by William Larue ( Figure 5 View FIGURE 5 ) shortly after capture clearly shows the presence of two, serrated spines on left posttemporal bone. This observation is confirmed by the radiograph which shows the posttemporal to be bifurcated posteriorly into the bases of two spines.
This configuration, with two posttemporal spines, is different from P. fraseri (Fowler 1934) , P. tridentifer ( McCulloch 1914) , P. inermis ( Borodulina 1981) , P. parini ( Borodulina 1979) , P spinifer ( Borodulina 1979) , P. soelae (Harold 1994) , P. stereope ( Jordan & Starks 1904), P. spinosus ( Günter 1887) , P. elongatus ( Borodulina 1979) , P. paxtoni ( Harold 1989) which have three and P. latirastrus which has one. Five species of the P. spinosus species group have posttemporals with two spines: P. asper (Harold 1994) , P. indicus ( Schultz 1961) , P. limatulus , P. nuttingi ( Gilbert 1905) and P. oluolus ( Baird 1971) . Polyipnus laruei differs from P. indicus , P. limatulus , P. nuttingi and P. oluolus by the presence of spines on ACB photophore scales, and by the lack of the second and third OVB photophores.
P. laruei is most similar to P. asper in regards to the structure of the preopercle, which has one long ventral spine, anteriorly curved, directed anteroventrally, and one short dorsal spine, laterally directed. These two species also share the presence of a compact ACC photophore cluster. The new species is distinguished from P. asper by a greater number of gill rakers on the first arch (25–26 versus 17–20, respectively), a much larger eye (eye diameter 22.4 % SL, as compared with 13.7–18.4), a lower vertebral count (32 versus 33), a smaller number of ACB photophores (6 in P. laruei and 8–10 in P. asper ), and ACA separated from ACB photophores. The comparison of meristic characters and photophores between holotype Polyipnus laruei and Polyipnus asper are given in Table 4. Polyipnus laruei also bears some similarities with P. oluolus from the Marshall Islands, in particular a large eye (22.4 % SL in P. laruei and 22.0 % SL in P. oluolus , versus approximatively 13–19 % SL in other Polyipnus spp.), and the presence of only 6 ACB photophores (4 in P. fraseri , and 7–13 in the remaining species in the genus).
The single specimen of Polyipnus laruei has some features that suggest paedomorphosis. These features include a large eye (22.4 % SL eye diameter) and a low count of ACB photophores (6). A relatively large eye (20.3 % SL) in P. fraseri (40.4 mm SL, holotype and 32.4 mm SL non-type) and low counts of ACB photophores were interpreted by Harold (1990b) to be paedomorphic. Among other Polyipnus species, only P. fraseri has a lower value (4 ACB) and only one other species has the same value as P. laruei ( P. oluolus ; 6 ACB). The remaining species have a range of ACB counts from 7 to 14. In sternoptychids individual photophores are added to the ventral clusters during development by a budding process ( Ahlstrom et al., 1984), which continues throughout most of the juvenile stage, although in P. laruei , P. fraseri and P. oluolus the evidence suggests that the rate of ACB photophore addition is retarded. Juvenilized morphology has been reported in other fishes, with examples from shallow water marine (Gobiiformes, Schindleria ; Johnson & Brothers, 1993), deep-sea ( Stomiiformes , Cyclothone ; Ahlstrom et al., 1984), and freshwater environments ( Cypriniformes , Paedocypris spp.; Kottelat et al., 2006) and is probably common throughout much of organismal diversity ( Gould, 1977).

| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |