
Cryptocellus luisedieri, Botero-Trujillo, Ricardo & Pérez, Gustavo A., 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.186535 |
|
publication LSID |
lsid:zoobank.org:pub:FC1E454F-BDEB-4A6A-B4FE-1F99CB2441C3 |
|
DOI |
https://doi.org/10.5281/zenodo.6213845 |
|
persistent identifier |
https://treatment.plazi.org/id/D65187E2-FFCA-FFD5-FF36-D46EFE817FC7 |
|
treatment provided by |
Plazi |
|
scientific name |
Cryptocellus luisedieri |
| status |
sp. nov. |
Cryptocellus luisedieri View in CoL sp. nov.
Figures 1–11 View FIGURE 1 View FIGURE 2 View FIGURES 3 – 6 View FIGURES 7 – 11
Type material. Holotype: Adult male from forest in the high basin of the rivers Rumiyaco and Ranchería, Kofán Territory at 00°03’N 77°13’W and 1430 m elevation, Ipiales Municipality, Nariño Department, Colombia; collected in pitfall trap, October 1998, E. González. Deposited in the “Instituto de Investigación de Recursos Biológicos Alexander von Humboldt”, Villa de Leyva, Colombia (IAvH-E 100812). The specimen, preserved in 70% ethanol, has the abdomen and both legs IV separated from the prosoma; the dissected copulatory apparatus is stored in a microvial with the specimen, as recommended by Platnick (2002). No additional specimens are known.
Distribution. Known only from the type locality ( Fig. 1 View FIGURE 1 ).
Etymology. Patronym dedicated to Luis Edier Franco (Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Villa de Leyva, Colombia), who has collected many new and interesting arachnid species in several Colombian localities, and has repeatedly helped us examine abundant material deposited at his institution.
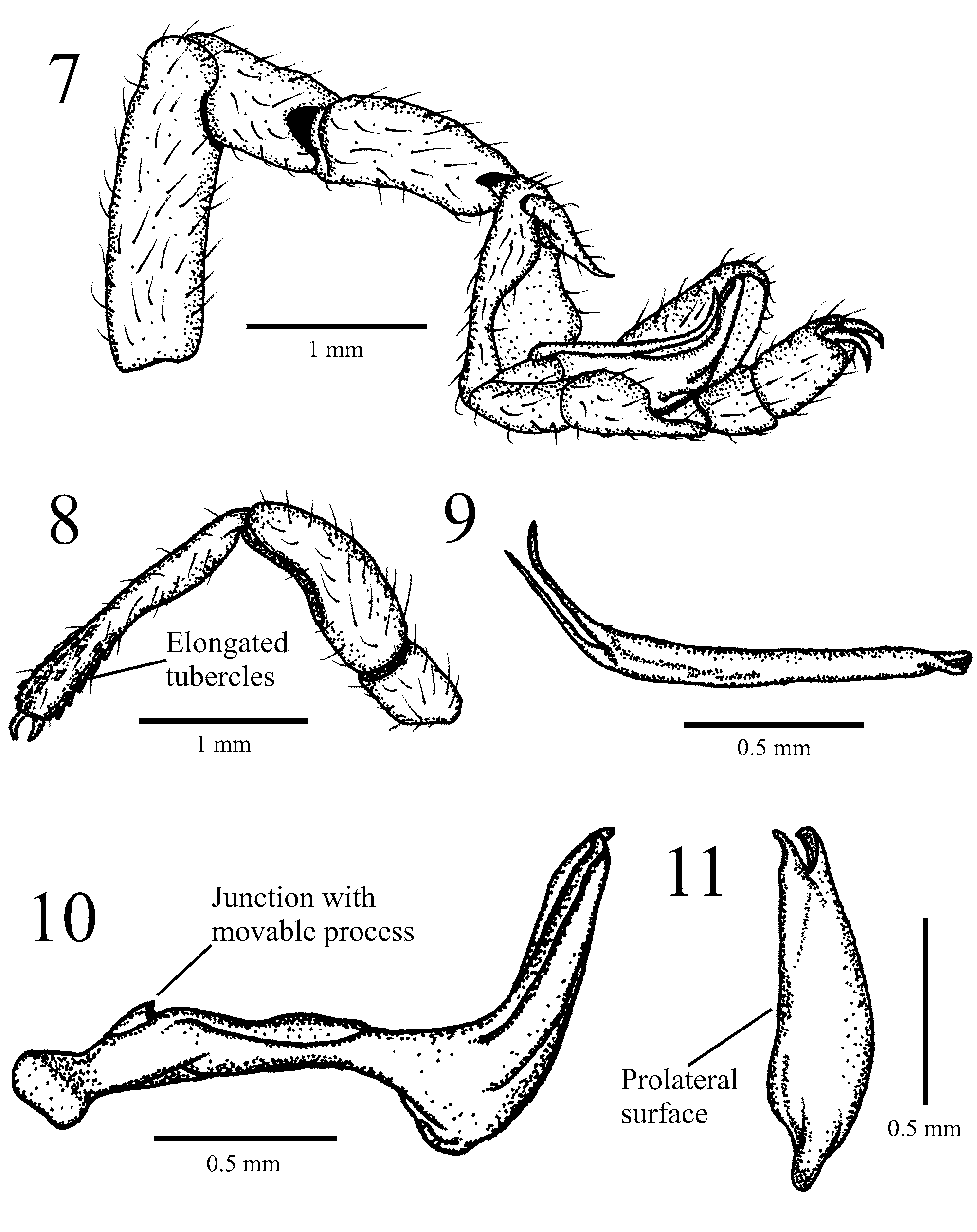
Diagnosis (based on male only). In having deep cuticular pits on carapace and abdomen ( Figs. 3, 5–6 View FIGURES 3 – 6 ) Cryptocellus luisedieri sp. nov. resembles Cryptocellus bordoni ( Dumitresco & Javara-Balş, 1977) from Venezuela, Cryptocellus brignolii Cokendolpher, 2000 from Suriname, C. magnus and C. narino from Colombia, and Cryptocellus pseudocellatus Roewer, 1952 from Peru. From the first four species, C. luisedieri sp. nov. differs in the highly distinctive shape of the copulatory apparatus: fixed process basally thin, straight, with well developed ventro-subdistal pronouncement, strongly curved distally, and bifurcated apically ( Figs. 7, 10–11 View FIGURES 7 – 11 ); movable process thin, predominantly straight but curved distally, and markedly bifid apically ( Figs. 7, 9 View FIGURES 7 – 11 ). Although the male of C. pseudocellatus and the female of the new species are unknown, they can be distinguished on the basis of the elongated and elevated tubercles on the distal half of the palpal tibia, present in C. luisedieri sp. nov. ( Fig. 8 View FIGURES 7 – 11 ) but absent in C. pseudocellatus . The new species also differs from C. brignolii , C. magnus and C. narino since the former species lacks cuticular pits on cucullus ( Fig. 4 View FIGURES 3 – 6 ), from C. narino by the lack of scale-like setae on legs, and from C. bordoni and C. brignolii in the unnotched pygidium.
Female. Unknown.
Male. Coloration: Body and appendages predominantly dark-red ( Fig. 2 View FIGURE 2 ), except ventral surface of abdomen brownish; carapace with pale, yellowish areas located at margins between legs I and II, suggesting corresponding position of ocular areas; abdominal articular membranes yellow.
Setation: Carapace, cucullus, abdomen and legs densely covered with fine translucent setae uniformly distributed throughout ( Figs. 3–7 View FIGURES 3 – 6 View FIGURES 7 – 11 ).
Carapace: Lateral margins not parallel (carapace narrowing anteriorly); slightly longer than wide, widest at level of leg III ( Fig. 3 View FIGURES 3 – 6 ); apparently without any vestige of ocelli except for above-mentioned pale regions (coated with setae); without granulation; shallow, median, longitudinal furrow present ( Fig. 3 View FIGURES 3 – 6 ), beginning at level of junction between coxae I and II, ending at level of leg III. Cuticle with deep pits containing tubercles, arranged as follows ( Fig. 3 View FIGURES 3 – 6 ): median longitudinal row of about nine, with eighth (counting from anterior end) noticeably wider and deeper than others; posterior margin with seven on each side of midline; short oblique submarginal rows each made up of four.
Cucullus: Wider than long, noticeably widest anteriorly; with conspicuous granulation anteriorly and laterally; devoid of pits; with complete, shallow, longitudinal furrow ( Fig. 4 View FIGURES 3 – 6 ).
Chelicerae: Movable fingers twice length of fixed fingers; left and right movable fingers armed with seven and eight rounded teeth respectively, distally decreasing in size, the two most basal teeth enlarged and forming a bicuspid; both fixed fingers with six rounded teeth similar in size, except second most basal tooth of both chelicerae and most distal on left chelicera larger.
Sternal region: Coxae I almost meeting tritosternum; coxae II, III meeting entirely, coxae IV meeting anteriorly; coxae II wider than others.
Abdomen: Longer than wide, widest at posterior margin of tergite XII ( Fig. 5 View FIGURES 3 – 6 ); devoid of granules; median plates of tergites XI–XIII with paired lateral depressions beside median elevation; all median plates slightly wider than long ( Fig. 5 View FIGURES 3 – 6 ). Cuticle with deep pits containing tubercles, arranged as follows ( Figs. 5–6 View FIGURES 3 – 6 ): tergite XI median plate with about seven along anterior margin, eight on posterior margin, four on each lateral margin; tergite XII median plate with about nine on posterior margin, row of five located at each side of median elevation; tergite XIII median plate with about four on each side of median elevation; internal margin of lateral plates each with about five on tergites XI–XII, three on tergite XIII; sternites X–XIII with about 20 along lateral margins, not precisely in longitudinal arrangement, and with conspicuous elongated pit-like excavations on anterior margin of sternites XI–XIII in paramedian position. Basal segment of pygidium unnotched on either dorsal or ventral posterior borders.
Pedipalps: With few granules on coxa and trochanters; femur curved and considerably widened; distal half of tibia covered with elongate, elevated tubercles ( Fig. 8 View FIGURES 7 – 11 ); both claws without teeth.
Legs: Legs I and II with abundant spine-like granules dispersed on all segments, mainly on ventral surface; legs III and IV almost devoid of granules; legs II bulkier than others ( Fig. 2 View FIGURE 2 ); trochanter IV unmodified; tarsal claws strongly curved.
Copulatory apparatus: As in Figures 7, 9–11 View FIGURES 7 – 11 ; fixed process basally thin, straight, with well developed ventro-subdistal pronouncement, strongly curved distally, bifurcate apically; movable process thin, predominantly straight but curved distally, markedly bifid apically.
Measurements (in mm): Total body length, excluding pygidium 7.67; cucullus 1.10 long, greatest width 1.71; carapace 2.38 long, 2.05 wide between legs II–III; abdomen 4.19 long (excluding pygidium), 2.90 wide near posterior margin of tergite XII (where widest); pedipalp femur 1.24 long, greatest diameter 0.52; pedipalp tibia 1.81 long, greatest diameter 0.24; femur I 1.43 long, 0.52 in diameter; femur II 2.57 long, 0.82 in diameter.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |