
Trechus balticus, Schmidt, Joachim & Faille, Arnaud, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4028.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:CB2162F6-6C18-445E-BAF1-D3B8C7DCF181 |
|
DOI |
https://doi.org/10.5281/zenodo.6100098 |
|
persistent identifier |
https://treatment.plazi.org/id/D40A87EA-2005-FF81-5782-D2F1502495C5 |
|
treatment provided by |
Plazi |
|
scientific name |
Trechus balticus |
| status |
sp. nov. |
Trechus balticus View in CoL sp. n.
Figs. 1–11 View FIGURES 1 – 2 View FIGURES 3 – 6 View FIGURES 7 – 10 View FIGURES 11 .
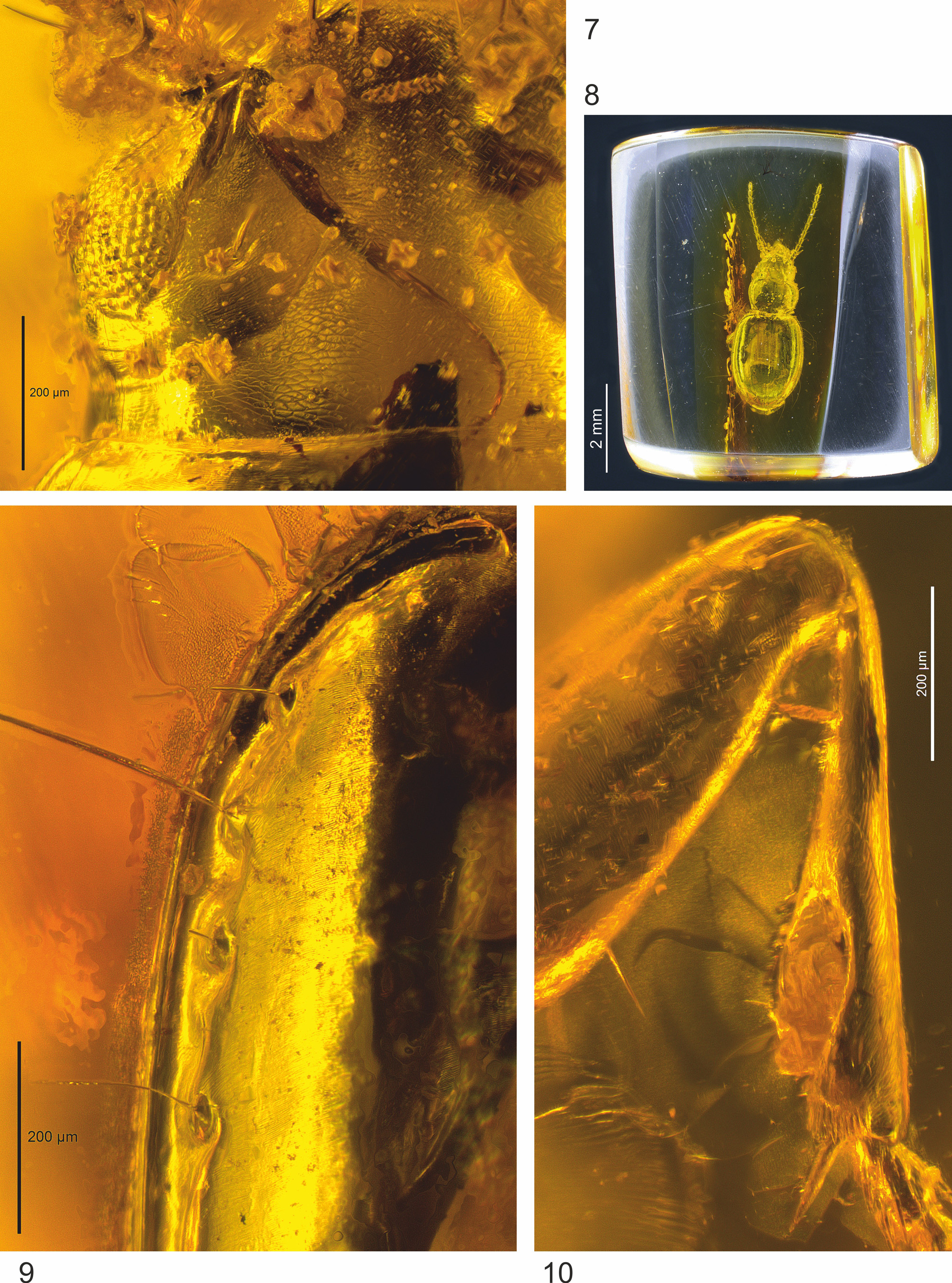
Holotype. Female in Baltic amber; size of piece 7.9 x 6.5 x 4.5 mm, embedded in synthetic resin ( Hoffeins 2001), No. 1310-5a in the collection of Christel and Hans Werner Hoffeins, Hamburg, Germany ( CCHH) (later to be deposited at the Senckenberg Deutsches Entomologisches Institut, Müncheberg, Germany) ( Fig. 8 View FIGURES 7 – 10 ).
Preservation status. Most external parts of the beetle body are well visible under optical microscope; the mouth parts are surrounded by milky coating and thus, mandibles (only anterior margin is visible), maxillae and mentum cannot be examined using light microscopy.
Syninclusions. A part of the amber piece that includes one specimen of Diptera and two plant trichomes, was separated and deposited under the number 1310-5b in CCHH.
Description. Body length: 3.6 mm.
Colour: Surface of body apparently blackish brown (artifact?), shiny; palpi, antennae and legs not distinctly lightened.
Microsculpture: Disc of head with rather deeply engraved, large and slightly transverse micro-meshes ( Fig. 7 View FIGURES 7 – 10 ). Disc of pronotum and surface of elytral intervals with very fine markedly transverse meshes, appearing finely fasciated even under high magnification (see Fig. 9 View FIGURES 7 – 10 : magnification 160 x).
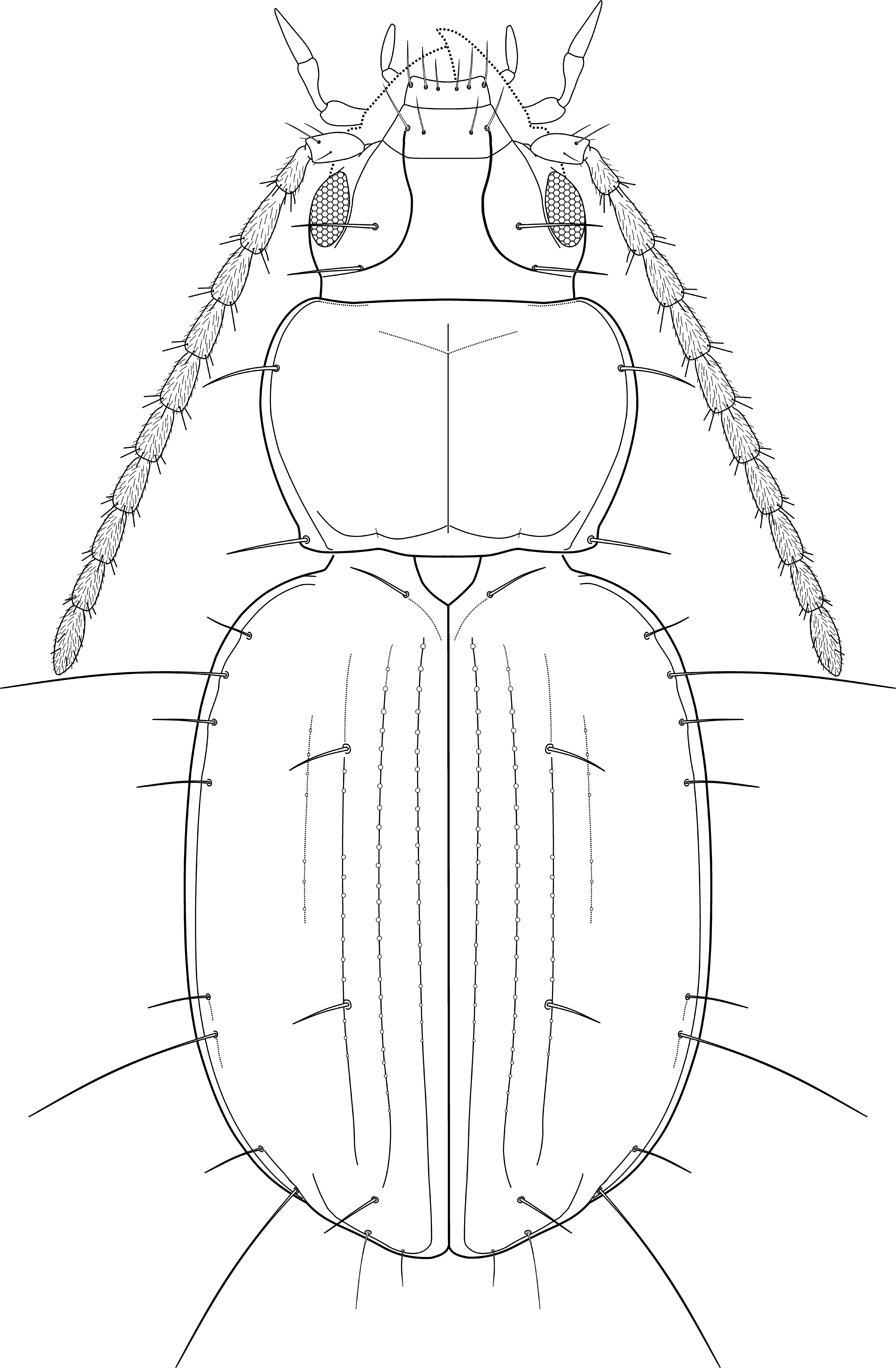
Head: Short and robust, notably broad near base. Mandibles relatively short. Labrum with three pairs of setae near moderately concave apical margin. Clypeus with two pairs of setae in normal position. Shape of maxillary and labial palpi as in Trechus sensu stricto. Eyes moderately small and slightly protruded; temples convex and rather short (length of eye/length of temple = 0.45), smooth. Frons and supraorbital areas markedly convex, with supraorbital furrows deep, complete, and uniformly bent on disc; two supraorbital setae present in normal position for Trechus . Antennae moderately short, with the tip of the eighth antennomere extended to the pronotal basal margin.
Prothorax: Pronotum 1.37 times broader than head, markedly transverse (width/length = 1.47), with sides evenly rounded in anterior 5/6 and emarginate at laterobasal angles, broadest somewhat anterior of middle, moderately narrowed towards base (width of pronotum/width of pronotal base = 1.3); basal margin 1.1 times broader than apical margin. Disk convex, smooth. Anterior margin very finely beaded in outer third, smooth in middle, almost straight, with apical angles slightly protruded and rounded. Posterior margin not beaded, moderately convex in middle, distinctly incised slightly before outer quarters, and again convexly rounded towards laterobasal angles ( Fig. 11 View FIGURES 11 ); latter relatively small, slightly shifted anteriad, slightly obtuse, not protruded laterally. Median longitudinal impression distinct, not deeper near base, disappearing near apex; anterior transverse impression very fine, smooth; posterior transverse impression and laterobasal foveae linear, smooth, pronotal base indistinctly flattened at laterobasal foveae. Lateral gutter relatively small throughout, slightly widened towards base, smooth. Both lateral and laterobasal setae present, with the lateral seta located at apical third of pronotum. Proepisternum smooth, prosternal process without margin.
Pterothorax: Elytra convex on disc ( Figs. 3, 4 View FIGURES 3 – 6 ), glabrous beside normal setation, in dorsal view relatively short, ovate (length/width = 1.25), with sides moderately convex ( Figs. 1 View FIGURES 1 – 2 , 11 View FIGURES 11 ), widest near their midlength, distinctly wider than pronotum (width of elytra/width of pronotum = 1.40). Humeral angles fully rounded, basal groove absent. Parascutellar stria finely developed, parascutellar seta present. First stria almost complete but disappearing at base; second and third striae somewhat more finely impressed than first, disappearing at apex; fourth stria indistinct, indicated and only barely visible at anterior half of elytra; outer discal striae absent; striae 1–4 finely punctate apart from the apical quarter of elytra. Each elytron with two discal setae in third stria, and with preapical seta, situated at apical tenth of elytra, almost equally as close to the apical margin as to the suture of elytra ( Figs. 6 View FIGURES 3 – 6 , 11 View FIGURES 11 ). Recurrent stria almost straight and relatively short, extending towards the reduced fifth stria anteriorly; front end of recurrent stria situated slightly anterior of the level of the preapical seta ( Figs. 6 View FIGURES 3 – 6 , 11 View FIGURES 11 ). Setae of umbilicate humeral series close to the elytral margin, with anterior three setae equidistant to margin and fourth seta located slightly further ( Figs. 9 View FIGURES 7 – 10 , 11 View FIGURES 11 ). The two setae of the medial group of the umbilicate series far removed from the preapical group, ca. twice the distance between the two setae of the preapical group ( Fig. 1 View FIGURES 1 – 2 , 11 View FIGURES 11 ). Epipleura long, nearly reaching the apex; metepisternum short, smooth, with outer margin only 1.15 times longer than anterior margin ( Fig. 5 View FIGURES 3 – 6 ).
Abdomen: All sternites smooth beside normal setation; sternites V–VI each with one, sternite VII with two (female!) pairs of setae near apical margin.
Legs: Averaged in length and form for Trechus ( Figs. 3, 4 View FIGURES 3 – 6 ), all femora unmodified; protibia straight and moderately dilated towards apex, without groove on outer surface ( Fig. 10 View FIGURES 7 – 10 ).
Etymology. The new species is named after the origin of the fossil species in the Baltic amber forests.
Species group recognition and relationships. Tribe: Trechus balticus sp. n. is here treated as a species of the tribe Trechini based on the combination of the following characters: frontal furrows semicircularly separating well developed supraorbital region from frons and neck; elytral sutural stria recurrent at apex, forming a deep carina directed towards the end of the fifth stria; apical segment of maxillary palpus as long as the penultimate, at base almost as broad as apex of penultimate segment, and at apex regularly conically pointed (the latter character distinguishes the fossil species also from the Trechini subtribes Aepina and Perileptina).
Genus: Trechus balticus sp. n. is a representative of the genus Trechus sensu lato based on the combination of the following characters: Mandibles normal, eyes glabrous, not markedly protruded, two supraorbital setae present. Pronotum glabrous, with one anterolateral and one laterobasal setae each side. Elytra glabrous, with basal groove absent, and with two discal setae; recurrent sutural stria directed towards the end of the fifth stria. The preapical seta is situated approximately at same distance to the apical margin as to the suture of elytra. The setae of humeral group are situated close to the elytral margin, with anterior three setae equidistant from margin; the medial group is well separated from the preapical group. Apical portion of protibia glabrous on external surface.
Group of Trechus sensu stricto: Molecular-systematic studies have shown that Trechus sensu lato is a nonmonophyletic assemblage ( Faille et al. 2010, 2011, 2013). E.g., T. bastropi Schmidt, 2009 from the Tibetan dacatraianus species group was identified sister to the genus Doderotrechus Vigna Taglianti, 1968 from the southeastern Alps ( Faille et al. 2013). However, species of the T. dacatraianus group are easy to distinguish from the fossil species as well as from the main Trechus clade by distinctly pubescent temples.
Another enlightening result coming from the molecular phylogenetic studies is the phylogenetic position of the species group of T. schaufussi Putzeys, 1870 endemic to the Iberian Peninsula. This lineage groups together with the genera Paraphaenops Jeannel, 1916 , Sardaphaenops Cerruti & Henrot, 1956 , and Typhlotrechus J. Müller, 1913 , comprising highly specialized hypogean species from the Mediterranean region ( Faille et al. 2013). Using external characters, species of the T. schaufussi group differ from the fossil species, above all, by the shape of the pronotum with median impression markedly deepened posteriorly, with basal transverse impression wide and not sharply defined, and with laterobasal fovea very large. The same characters distinguish also T. rubens (Fabricius, 1792) and T. ovatus Putzeys, 1846 from the fossil species. Both the former taxa are considered representatives of Trechus sensu stricto by previous authors too. However, in the molecular phylogenetic analysis these species group with the Palaearctic taxon Epaphius Stephens, 1827 ( Faille et al. 2013) . Species of the Epaphius group sensu Jeannel (1927) can easily be distinguished from the fossil species by the different position of the elytral preapical seta which is markedly shifted anteriorly, and, therefore, missing apical triangle.
A pronotal shape similar to the fossil T. balticus sp. n., with posterior transverse impression and laterobasal foveae linear and sharply limited towards the convex disc, is also observed in several Palaearctic species of the main Trechus clade sensu Faille et al. (2013), e.g., in the type species of the genus Trechus , T. quadristriatus (Schrank, 1781) , in the Southeast Asian T. indicus group sensu Jeannel (1927), and in the highly diverse Himalayan-Tibetan T. thibetanus group sensu Schmidt (2009). But this character is also present in Nearctic species of the genus (e.g. T. hydropicus G. Horn, 1883 , or species of the subgenus Microtrechus Jeannel, 1927 ) ( Barr 1962). Microtrechus is distinguished from the main Trechus clade sensu Faille et al. (2013), e.g., by the presence of only one dilated male protarsomere. However, since the fossil specimen is a female the character state remains open in this respect.
Consequently, the fossil T. balticus sp. n. may represent an extinct species of one of these species groups. A comprehensive enlargement of the taxon sampling of the molecular studies is needed for a better understanding of morphological character evolution within the mega-diverse Trechus sensu lato. On the other hand, additional Trechus fossils should be carefully investigated to find further evidence for the systematic positions of representatives of the Eocene Trechini fauna, and additional characters have to be investigated, e.g. male genital characters. The latter might be possible using X-ray MicroCT techniques on amber specimens (e.g., Perreau & Tafforeau 2011, Kehlmaier et al. 2014).
Differential diagnosis. The new fossil species is easily distinguishable from the Baltic amber fossil Trechoides fasciatus with uncertain systematic position due to the much larger body (body length of Trechus balticus sp. n. approx. two times of Trechoides fasciatus ).
To date, no additional Trechini species are known from Baltic amber and other Eocene deposits identified at the species level. Therefore, we present a differential diagnosis with respect to the modern Trechus species.
As already discussed previously, based on characters of the pronotal shape and elytral chaetotaxy, the new fossil species is similar to members of the T. quadristriatus and T. indicus species group sensu Jeannel (1927), including the species of the T. thibetanus group sensu Schmidt (2009). Trechus balticus sp. n. is easy to distinguish from most representatives of these groups, due to the distinct reduction of the elytral striation ( Fig. 11 View FIGURES 11 ), with only the first three striae slightly impressed on disc, the fourth stria only partly recognizable in middle, striae 5–7 absent, and with remains of the eighth stria slightly impressed at levels of the medial and subapical groups of the umbilical series. A similar reduction of the elytral striation is also developed in some species of the T. thibetanus group sensu Schmidt (2009). However, all species of that group except T. glabratus Schmidt, 2009 , have microsculpture pattern on elytra clearly isodiametric, while microsculpture is markedly transverse in T. balticus sp. n. In T. glabratus the eighth elytral stria is deeply impressed also between the medial and subapical groups of the umbilical series while it is absent in T. balticus sp. n.
The fossil species is also similar to the Western Nearctic Trechus sensu lato species ( Microtrechus and T. hydropicus group) with which it shares numerous characters, like a similar robust habitus, short elytra, long epipleura, only the first elytral striae present, microsculpture transverse. However, it differs by a more transverse pronotum and the lack of the protibial groove.
Biogeographical and ecological considerations. The genus Trechus sensu lato (approx. 900 modern species) is widely distributed in the Holarctic region, with more than 90% of the species occurring in the Palaearctic, and 69 species in the Nearctic ( Bousquet 2012); some moderately diverse species groups are also known from the mountains of East Africa. Therewith, the occurrence of Trechus in Baltic amber is in accordance with the modern distribution of the genus, as it is observed for most of the other ground beetle species described from Baltic amber at species level ( Bembidion Latreille, 1802 , Calathus Bonelli, 1810 , Dromius Bonelli, 1810 , Limodromus Motschulsky, 1850 , Loricera Latreille, 1802 ). Since the true relationships of the fossil species with respect to the modern lineages remain open (e.g., closer affinities to Palaearctic or Nearctic lineages?) no further biogeographical conclusion can be made at the current stage.
Because T. balticus View in CoL sp. n. has short metepisternae, it is highly probable that the hind wings of the fossil species are reduced to short stubs or absent. In Trechini View in CoL , shortened metepisternae and reduced hind wings generally indicate a strictly edaphic mode of life. It can thus be assumed that the specimen was trapped by the resin at the base of the producing tree or on the forest floor. This is in accordance with the conclusion of Larsson (1978), that arthropods from the forest floor represent an important portion of the Baltic amber specimens. For example, two other strictly edaphic ground beetle species, Dyschiriomimus stackelbergi View in CoL of the tribe Scaritini View in CoL , and Loricera electrica View in CoL of the tribe Loricerini , were described by Yablokov-Khnzorian (1960), and Klausnitzer (2003), respectively. With respect to additional Carabidae View in CoL taxa preserved in Baltic amber, it is uncertain whether they are trapped while moving on the tree (e.g., Lebiini View in CoL , Platynini View in CoL , Tachyina View in CoL ), running on the ground or, as a result of an accident while flying.
With respect to the extant fauna, most if not all wingless epigean Trechus View in CoL species are occurring exclusively in mountainous regions. This makes it very likely that also the fossil T. balticus View in CoL sp. n. was a montane species. With respect to the environment of the Baltic amber producing forests, which is considered to be situated in the paratropical to subtropical zone of the Eocene ( Weitschat & Wichard 2010), the assumption of the existence of montane habitats would explain why a number of additional ground beetle taxa adapted to temperate climates are known from this amber depositions (e.g., Agonum View in CoL , Amara View in CoL , Calathus View in CoL , Limodromus ). At least warm temperate climatic conditions are assumed for the many higher elevated areas within the vast region that was covered by Baltic amber forests, which stretched from the southern slopes of the Fennoscandian shield in the West to the Ural Mountains in the East and to the Ukrainian highlands in the South ( Weitschat & Wichard 2008, 2010).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |