
Careproctus phasma Gilbert 1896
|
publication ID |
https://doi.org/10.11646/zootaxa.4018.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:BDEF6499-7A88-496E-AB24-A82977F0D1A5 |
|
DOI |
https://doi.org/10.5281/zenodo.5618223 |
|
persistent identifier |
https://treatment.plazi.org/id/D33887A3-FFBE-FF96-FF3E-FE06FD2AFC8D |
|
treatment provided by |
Plazi |
|
scientific name |
Careproctus phasma Gilbert 1896 |
| status |
|
Careproctus phasma Gilbert 1896 View in CoL
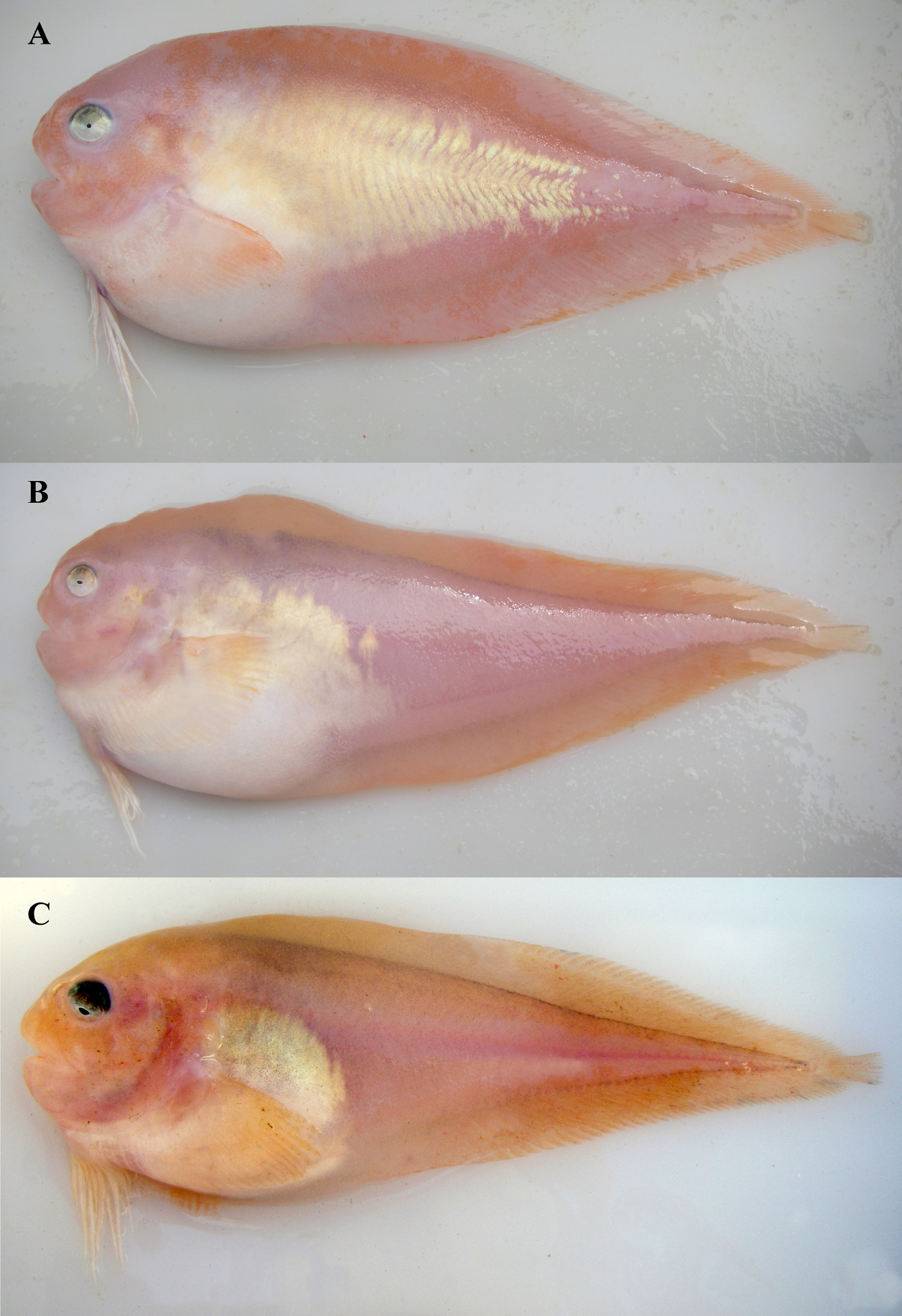
English common name: Spectral Snailfish Figures 3 View FIGURE 3 B, 4A, 5F, 6; Tables 1–3
Careproctus phasma Gilbert 1896:443 View in CoL . Type locality: Bristol Bay, Alaska, 56°50'00"N, 164°27'50"W.— Jordan & Evermann 1898:2132 (description).— Jordan & Gilbert 1899:478 (Bristol Bay).— Evermann & Goldsborough 1907:333 (Alaska).— Burke 1930:123, fig. 43 (description, key).— Taranetz 1937:137 (Bering Sea, in key).— Schmidt 1950:195 (brief description, Russia, possible misidentification, see below).— Böhlke 1953:136 ( type catalog).— Wilimovsky 1954:286 (checklist).— Wilimovsky 1958:78 (keys).— Isakson et al. 1971:669 (listed, probable misidentification, = C. scottae View in CoL ).— Quast & Hall, 1972:28 (checklist).— Fedorov 1973:66 (Bering Sea).— Allen & Smith 1988:67 (Alaska, “pink snailfish”, in part).— Pitruk 1990:38 (list, Bering Sea, Sea of Okhotsk from Schmidt, 1950).—Robins et al. 1991:44 (list).— Sheiko & Fedorov 2000:32 ( Russia).— Mecklenburg et al. 2002:618 (Alaska, in key, in part).— Parin et al. 2002:S113 (list).— Chernova et al. 2004:13 (checklist).— Nelson et al. 2004:124 (list).— Chernova 2005b:S7 (comparisons).— Kai et al. 2011a:153 (genetics, morphology, phylogenetics, as “BER3”).— Kai et al. 2011b:368 (fig. 1a, phylogenetics, as “BER3”).—Page et al. 2013:127 (list).
Lectotype. USNM 48064, ca. 85 mm, Bristol Bay, Alaska, Albatross station 3254, 56.83°N, 164.46°W, depth 84 m, poor condition, disintegrated.
Paralectotype. SU 3028, 72.3 mm, N. of Unimak I., Albatross station 3256, 56.3°N, 164.57°W, depth 90 m, 14 June 1890 ( Fig. 4 View FIGURE 4 A).
Additional material examined. A total of 119 specimens, not including the types above, 54.3–272.0 mm SL. See “Non-type material examined below.”
Diagnosis. Careproctus phasma is distinguished from all other species of Careproctus by the combination of cyt b and 16S rRNA sequences (“ARC1”, in part, and “BER3” of Kai et al. 2011a; Table 1), an anteriorly robust body that is either naked or covered with scattered cactus-like prickles, the absence of the postorbital pore, a moderate-sized pelvic disc, a lower pectoral-fin lobe longer than upper lobe, a light peritoneum and stomach, and lower counts of vertebrae and median fin rays. It is most similar to the new species C. lerikimae of the Beaufort Sea, from which it can be distinguished by its entirely pale coloration (vs. dusky body in C. lerikimae ), fewer vertebrae ( 55–60 in C. phasma vs. 59–63 in C. lerikimae ) and median fin rays (50–56 and 43–49 in C. phasma vs. dorsal 56–59 and anal 48–51 in C. lerikimae ), as well as its shorter maxilla, longer lower pectoral-fin lobe, and longer nasal tube. It is also similar to C. rastrinus and C. scottae from which it can be distinguished by the presence of the postorbital pore (absent in C. phasma ), larger pelvic disc (20.1–35.9 vs. 9.9–26.8 % HL), and lower counts of caudal vertebrae (44–50 vs. 48–56), dorsal-fin rays (50–56 vs. 54–63), and anal-fin rays (43–49 vs. 47–56). It is distinguished from C. spectrum by its smaller orbit (17.3–31.6 vs. 33.9–34.4 % HL).
Description. Body heavy and deep anteriorly, tapering posteriorly, moderately compressed; depth at pectoralfin base 82.9–136.6 (83.6) % HL. Head large, 22.4–29.7 (25.3) % SL, robust, dorsal profile rounded from nape to snout. Snout blunt, short, slightly projecting anterior to lower jaw, longer than orbit, 100.0–223.6 (128.9) % OL, 26.2–42.9 (31.7) % HL. Mouth terminal, small, horizontal; upper jaw 31.1–48.1 (44.3) % HL, maxilla extending to anterior part of orbit or mid-orbit, oral cleft extending to anterior rim of orbit; mandible 27.1–56.2 % HL. Premaxillary tooth plates matching mandibular tooth plates. Premaxillary and mandibular teeth simple with weak shoulders in 17–33 oblique rows of 7–9 teeth forming narrow bands. Diastema absent at symphysis of upper and lower jaws. Orbit 17.3–31.6 (24.6) % HL, dorsal margin at or just below dorsal contour of head, suborbital depth to upper jaw 12.1–18.2 (12.5) % HL, to lower jaw 24.1–33.6 (31.7) % HL; pupil round. Interorbital space broad, fleshy distance 19.6–52.5 (52.5) % HL, bony distance 14.0–32.5 (33.3) % HL, strongly convex. Nostril single, with well-developed tube at level with lower rim of orbit; nostril tube length 13.0–30.4 (15.6) % OL.
Pores of cephalic lateralis system of moderate size, pore pattern 2-5-7-2, chin pores paired. Interorbital pore absent. Gill opening small, 14.5–36.9 (34.4) % HL, upper margin at or just above level of dorsal rim of orbit, extending ventrally to just above the upper pectoral-fin ray to pectoral-fin ray 1–5 (above upper ray). Opercular flap rounded to slightly angular (rounded). Gill rakers 6–10 (Tables 2–3), short, blunt.
Dorsal-fin rays 50–56 (54; Tables 2–3), anterior dorsal lobe absent, anterior rays buried in tissue, tips of more posterior rays not exserted. Anteriormost dorsal-fin pterygiophore inserted between neural spines 3 and 4, bearing a single small ray. Predorsal length 24.0–32.8 (28.4) % SL. Anal-fin rays 43–49 (47; Tables 2–3), one to three analfin pterygiophores anterior to first haemal spine, each bearing a single ray, tips of rays not exserted. Anal-fin origin below vertebrae 12–13 (caudal vertebrae 1–2), preanal length 31.6–51.8 (41.1) % SL.
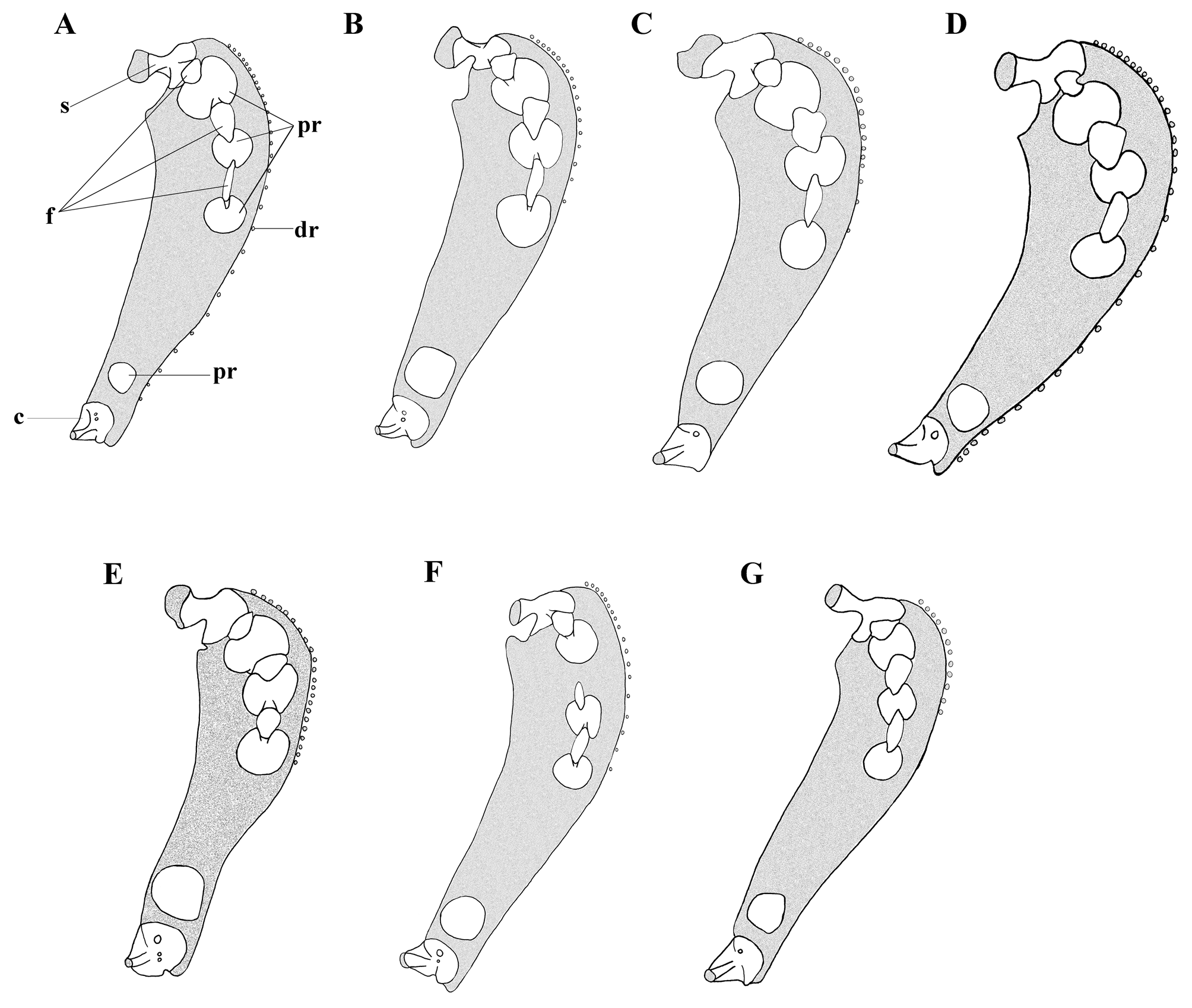
Pectoral fin deeply notched, with 27–35 rays (29; Tables 2–3). Upper lobe 51.5–86.6 (68.3) % HL, with 20–26 (21) rays, extending beyond anus short of anal-fin origin, shorter than lower lobe, dorsalmost rays lengthening to rays 5–7, more ventral rays gradually shortening to shortest ray of notch. Lower lobe moderately elongate, 60.3– 134.6 (70.5) % HL, with 6–9 rays (8; Table 2), extending beyond anus short of anal-fin origin; dorsal rays gradually lengthening to elongate rays 3–4, ventral rays gradually shortening to ventralmost ray near pectoral symphysis. Tips of rays of dorsal lobe 5–15% free of membrane, rays of lower lobe more strongly exserted, up to 30% free. Notch strong, rays in notch slightly more widely spaced than rays of lobes, more widely spaced ventrally. Uppermost pectoral-fin ray level with region between ventral rim of orbit and cleft. Insertion of lowermost pectoral-fin ray anteriorly placed, below area between tip of snout and anterior rim of orbit. Proximal pectoral radials four (3+1), small: radial 1 crescent shaped, radial 2 notched and distinctly hour-glass shaped, radial 3 crescent shaped, radial 4 round ( Fig. 5 View FIGURE 5 D). Interradial fenestrae three, extending between scapula and proximal radials 1–3: oval between scapula and radial 1; small, elongate, forming a notch on dorsal margin of radial 2, distant from radial 1; elongate, elliptical between radials 2 and 3; dorsoventrally elongate between radials 2 and 3. Scapula broadly T-shaped with equally sized robust arms and helve; coracoid with broad triangular head, large foramen, and short robust helve. Distal radials present at base of rays 2–18, ventralmost at level of proximal radial 3, more ventral rays articulating directly with pectoral cartilage.
Pelvic disc large, length 20.1–35.6 (27.2) % HL, round, slightly longer than wide, width 18.3–32.3 (30.6) % HL, anterior lobe weakly developed, slightly to strongly cupped, distance from tip of snout to pelvic disc 9.3–15.7 (10.2) % SL. Anus at a level just posterior to orbit, close behind pelvic disc; distance from snout to anus 13.7–27.8 (19.4) % SL, 54.4–102.8 % HL.
Principal caudal-fin rays 8–11, dorsal procurrent rays 1–2, ventral procurrent rays 0–2 (1–2 + 4/4–5/5–5/6 + 0– 2) (? + 5/5 +?). Caudal fin 32.9–54.8 % HL. Membrane of posterior dorsal-fin rays attached to caudal fin at shorter distance than anal-fin rays: dorsal-fin rays attached to caudal fin 31.5–65.3 % CL; anal-fin rays, 38.0–77.7 % CL. Depth at base of caudal fin 14.4–27.8 % CL.
Skin relatively thin, loose gelatinous layer particularly evident in juveniles, prickles reduced, in most dense region about 9 prickles in orbit length, or absent. Pyloric caeca 17–24, length about 28–37 % HL, left side of visceral cavity.
Vertebrae 55–60 (59), precaudal 10–12 (11), caudal 44–50 (48; Tables 2–3). Pleural ribs 2 or 3, anteriormost small when 3, others long and slender, present on vertebrae 8–10 or 9–10.
Coloration. Body pink and white and fins orangish pink in life ( Fig. 3 View FIGURE 3 B); head, dorsum from nape to caudal fin, and ventrum at anal-fin origin to caudal fin pink to marginally orange; isthmus, base of pectoral fin, and body posterior of gill slit to anal-fin origin white; area above belly silvery white (crystalline guanine) with orangish-pink highlights becoming obsolete posteriorly at about anal-fin origin; base and lower lobe of pectoral fin white; dorsal margin and distal portion of fin orangish pink; eye with dark dorsal margin, becoming brassy and whiter ventrally. Body and fins pale in preservation; base of fins beneath skin with pigment, showing faint line between fins and body. Peritoneum pale; orobranchial cavity pale; stomach, intestines, and pyloric caeca pale to dusky greenish; urogenital papilla pale.
Life history. The largest specimen examined was 272.0 mm (HUMZ 73075) of unknown sex. The smallest ripe female with yolked eggs was 102.9 mm; the largest, 176.7 mm. Three egg classes were found in ripe females: yolked eggs 3.5 mm, maturing white 1.8 mm, and immature < 1 mm. The smallest ripe male was 76.3 mm; the largest, 191 mm.
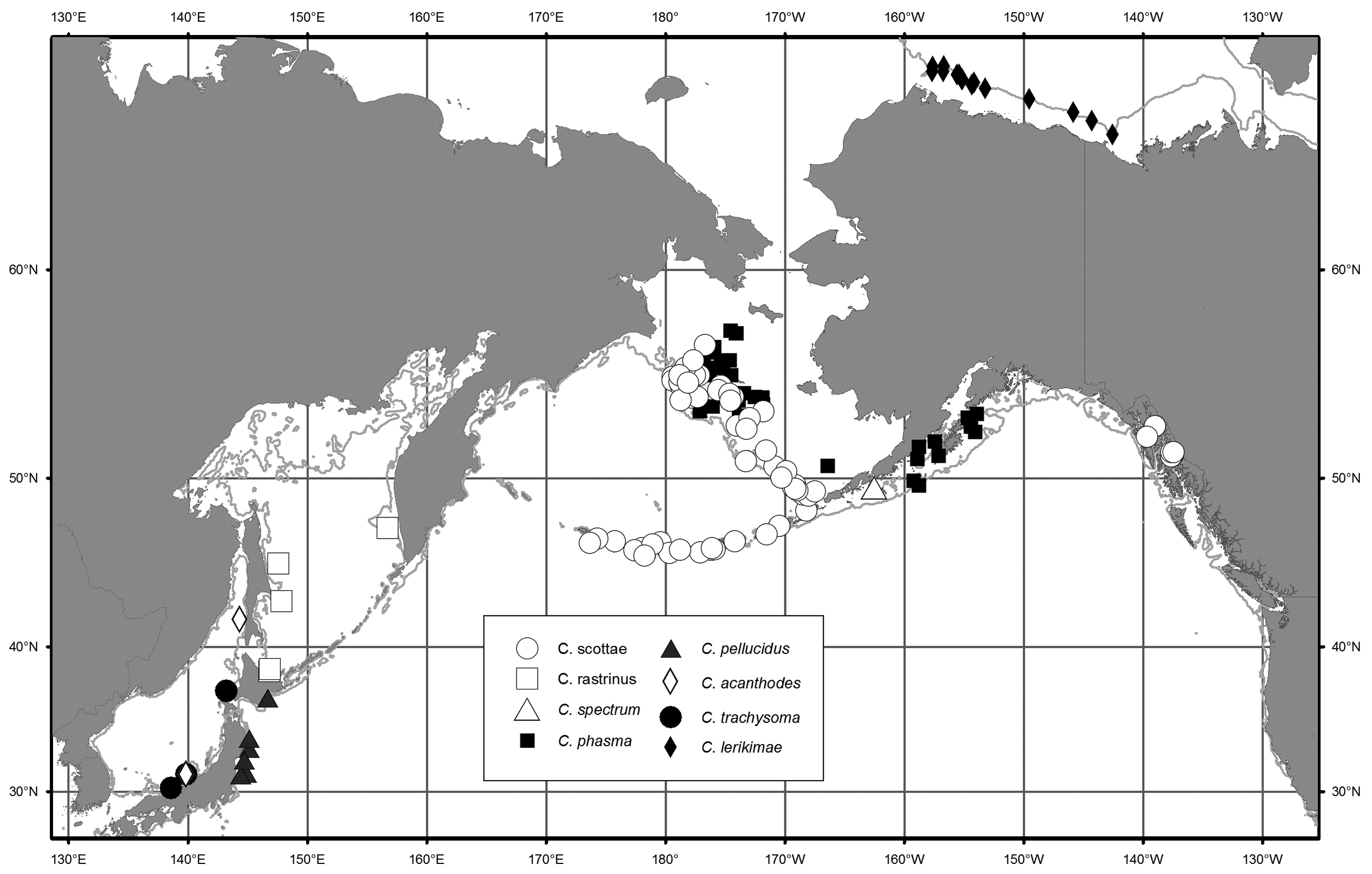
Distribution. Careproctus phasma has been collected from the continental shelf of the eastern Bering Sea and the northern Gulf of Alaska ( Fig. 6 View FIGURE 6 ). Collection depths range from 57 to 184 m.
Etymology. The specific epithet is taken from the Greek phasma (ϕάσΜα), meaning “specter”, “ghost”, or “apparition.”
Remarks. Gilbert (1896) described C. phasma from Bristol Bay in the Bering Sea and compared it to C. spectrum , known only from the type series. Based on his description of “the much larger sucking disk and the narrow gill-slit, the latter confined to area above base of pectorals”, Gilbert was clearly comparing C. phasma to C. gilberti Burke , which has a disk much smaller than the orbit and a large gill slit extending to about pectoral-fin ray 11 (Burke 1912). Careproctus gilberti Burke was described from 21 of the 26 syntypes of C. spectrum , and not the presently recognized types of C. spectrum .
Schmidt (1950) reported C. phasma from the Sea of Okhotsk based on two specimens. However morphological differences in the illustration provided by Mecklenburg et al. (2002), based on ZIN 29087 and reflecting features described by Schmidt (1950), indicate this species is unlikely to be C. phasma and may be an undescribed species. The pelvic disc is much longer than wide rather than nearly round, the pectoral fin is much longer (ca. 100% HL) than in our material (51.5–86.6% HL), and the rays of the lower pectoral-fin lobe are nearly entirely free. In addition, the collection depth of Schmidt’s specimens is 504 m, more than twice as deep as the relatively shallow-water C. phasma from the Bering Sea and Gulf of Alaska.
In the eastern Bering Sea, C. phasma is generally associated with the pool of cold water that extends from the north and in some years reaches the Alaska Peninsula (Stevenson & Lauth 2012). Temperatures recorded at depth during trawls in which C. phasma was caught ranged from -1.7 to 3.4°C (mean 0.3), in contrast with C. scottae , which was captured in temperatures of -1.2 to 4.7°C (mean 2.8). In the Gulf of Alaska, however, C. phasma was collected in temperatures ranging from 4.4 to 7°C; no temperature data was available for collections in the Gulf of Alaska of C. scottae .
In their phylogenetic analysis using cyt b and 16s data, Kai et al. (2011a) recovered a clade labeled as ARC1 as sister of the clade BER3 ( C. phasma ). “UW Uncataloged” was cataloged as UW 154442, a specimen of C. phasma from the northern Gulf of Alaska. Most similar to C. phasma , it differs from by two base pairs from nearly all others and by one base pair from C. lerikimae

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Careproctus phasma Gilbert 1896
| Orr, James Wilder, Kai, Yoshiaki & Nakabo, Tetsuji 2015 |
Careproctus phasma
| Kai 2011: 153 |
| Kai 2011: 368 |
| Chernova 2004: 13 |
| Nelson 2004: 124 |
| Mecklenburg 2002: 618 |
| Sheiko 2000: 32 |
| Pitruk 1990: 38 |
| Allen 1988: 67 |
| Fedorov 1973: 66 |
| Quast 1972: 28 |
| Isakson 1971: 669 |
| Wilimovsky 1958: 78 |
| Wilimovsky 1954: 286 |
| Bohlke 1953: 136 |
| Schmidt 1950: 195 |
| Taranetz 1937: 137 |
| Burke 1930: 123 |
| Evermann 1907: 333 |
| Jordan 1899: 478 |
| Gilbert 1896: 443 |