
Aberrantidrilus mihaljevici Vučković, Rodriguez & Kerovec, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4758.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:C6470FB8-3E44-45BA-9130-BC3EBD4EDE54 |
|
DOI |
https://doi.org/10.5281/zenodo.3812154 |
|
persistent identifier |
https://treatment.plazi.org/id/2E4DF7C5-6423-448C-A593-D73DB7B144EB |
|
taxon LSID |
lsid:zoobank.org:act:2E4DF7C5-6423-448C-A593-D73DB7B144EB |
|
treatment provided by |
Plazi |
|
scientific name |
Aberrantidrilus mihaljevici Vučković, Rodriguez & Kerovec |
| status |
sp. nov. |
Aberrantidrilus mihaljevici Vučković, Rodriguez & Kerovec n. sp.
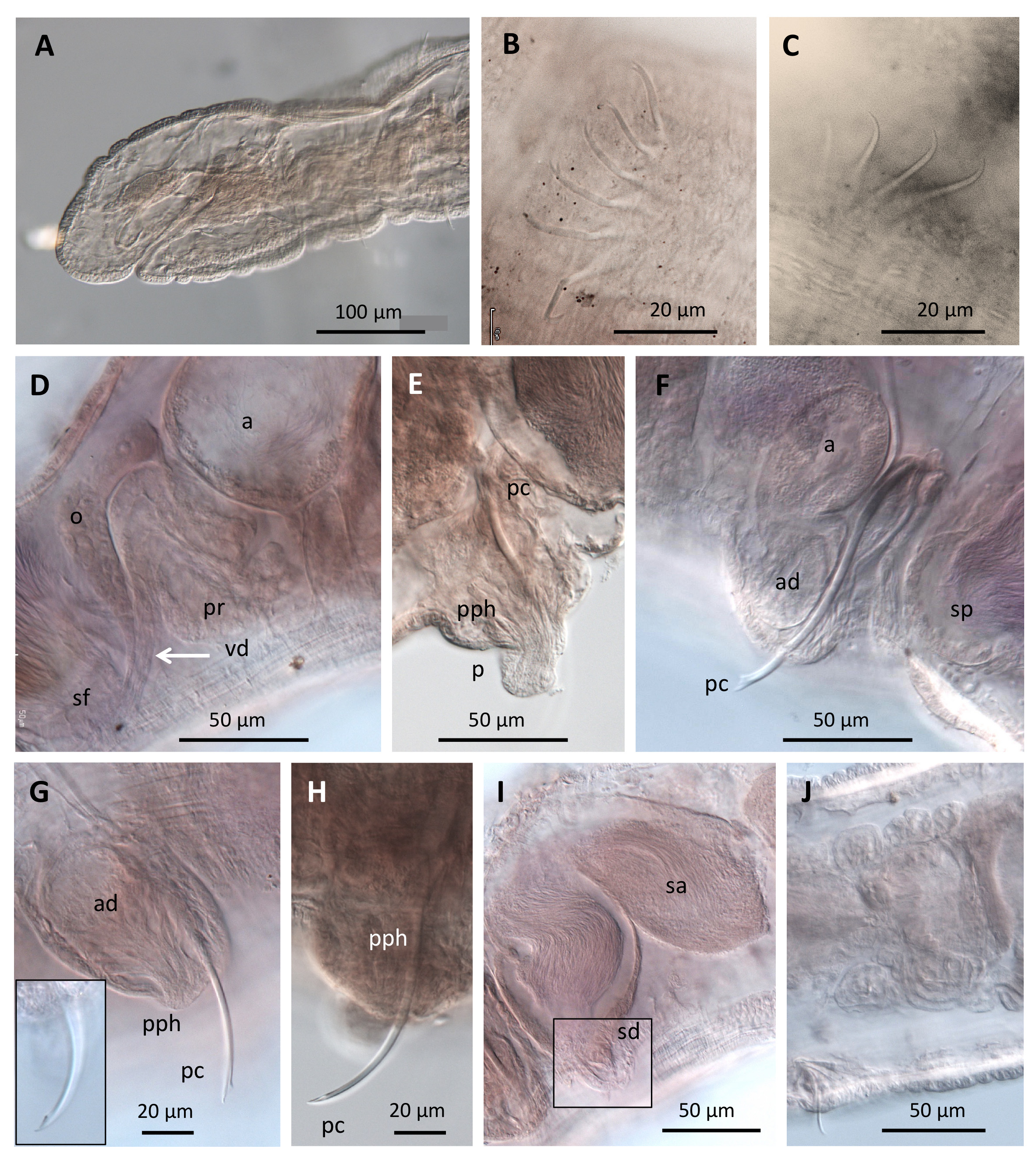
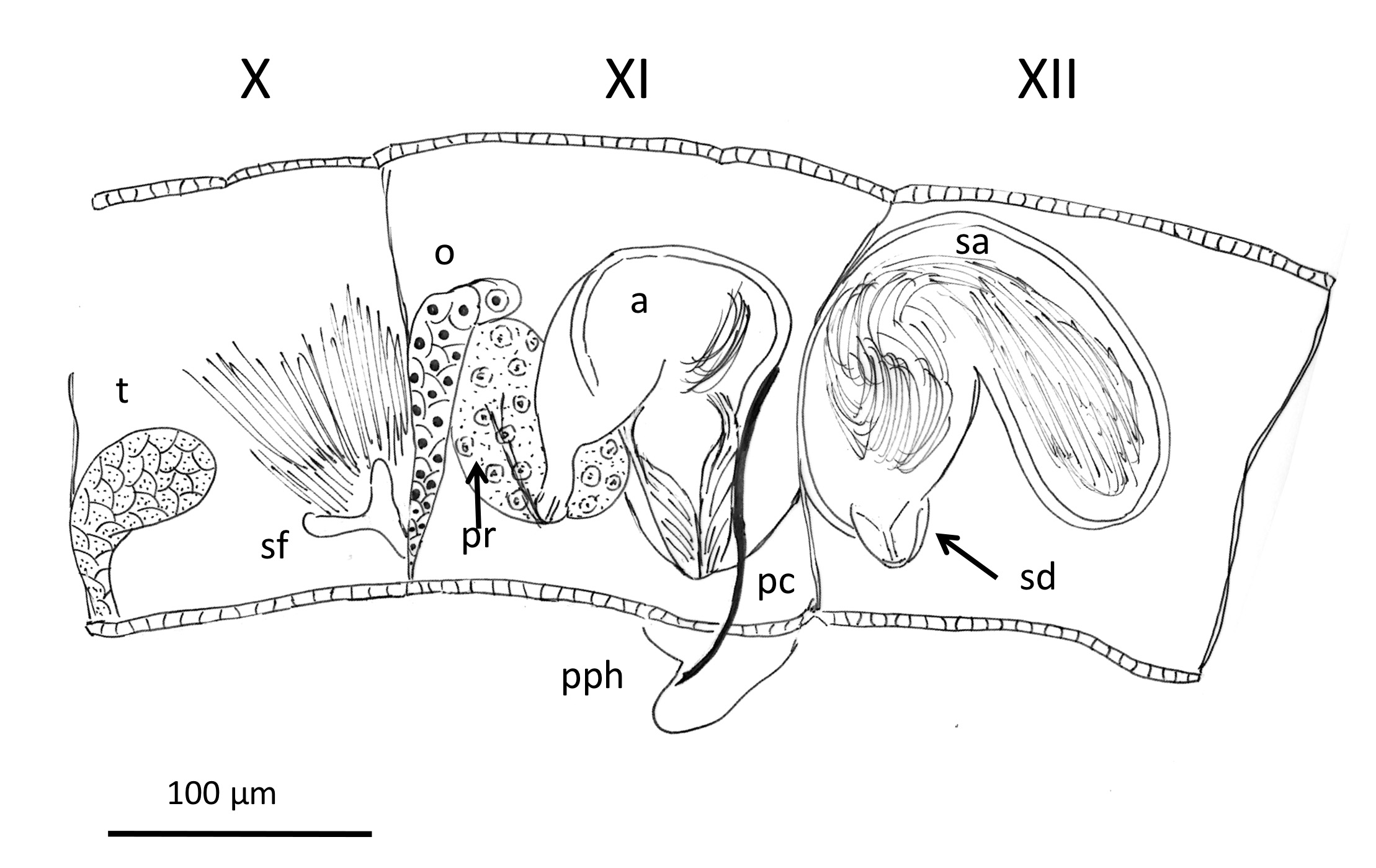
( Figures 2 View FIGURE 2 , 3 View FIGURE 3 )
Holotype. MNCN16.03 View Materials /3103, one whole-mounted individual, stained with hematoxylin and mounted in Canada balsam.
Paratype. MNCN16.03 View Materials /3104, one whole-mounted individual,stained in hematoxylin and mounted in Canada balsam, from type locality .
Type locality. In the tufa barrier of Roški slap, Croatia. April 28, 2014. Coordinates: 43°54’12.1”N, 15°58’31.3”E (43.90336 N, 15.975348 E). Collector: Zlatko Mihaljević.
Etymology. The species is named to honor its collector, Dr. Zlatko Mihaljević, professor of Zoology at the University of Zagreb, for his interest and contribution to the study of the biota in tufa barriers of Croatia.
Description. Specimens posteriorly incomplete, number of segments 14 (paratype) and 33 (holotype). Length of the longest worm 3 mm; maximum diameter 234 and 248 μm in segments IX and XI, respectively. Prostomium 71 μm long and 80 μm wide at its base ( Fig. 2A View FIGURE 2 ). Body wall consisting of a thin cuticule, an epidermal layer (4–7 μm thick) and musculature (5–6 μm thick). Clitellum not raised, formed by epithelial glands in transverse lines, from segment X behind the chaetal bundles to the anterior third of XII. One pair of simple spermathecal pores in the most anterior part of segment XII, opening in line with ventral chaetae. One pair of male pores in segment XI on porophores (36 μm high, 61 μm wide), in line with ventral chaetae. Dorsal and ventral chaetae bifid in anteclitellar segments, simple-pointed from XIV; in XII and XIII chaetae simple-pointed in dorsal bundles and bifid in ventral bundles ( Figs. 2B,C View FIGURE 2 ). Anterior segments with (3)4–7 chaetae per bundle and postclitellar segments with 5–7; chaetae usually more numerous in II and in the posterior segments ( Table 1). Length of chaetae gradually increasing from II to VI (34–44 μm), with distal tooth about the same length as proximal, and nodulus about median. Chaetal bundles situated in the posterior third of the segment. Penial chaetae occurring singly in segment XI, sigmoid, the tip grooved like the tip of a hypodermic needle ( Fig. 2H View FIGURE 2 ), or frayed, spatulate tip (only seen when mounted in Canada balsam) ( Fig. 2F, G View FIGURE 2 ), and oriented anteriad; length 117–124 μm, 2.7–3.2 times the length of somatic chaetae, and thickness 4 μm.
No coelomocytes observed within the coelomic cavity. Pharyngeal glands in segment III dorsal to the pharynx, and from IV–VI both dorsal and ventral to the oesophagus. Chloragogen cells from segment VI, backwards. The beginning of the intestine well-defined by a dilatation of the gut in VIII. First nephridia in 6/7 ( Fig. 2J View FIGURE 2 ), and additional nephridia observed in some posterior segments.
One pair testes in segment X and one pair ovaries in segment XI ( Fig. 3 View FIGURE 3 ). Anterior sperm sacs extending to seg- ment IX and posteriorly to XII, ovisac to XIII containing yolk (one oocyte also visible in the holotype). Male ducts paired. Vas deferens (diameter 6–8 μm), ciliated, shorter than the atrium ( Fig. 2D View FIGURE 2 ), junction with atrium not clearly determined, although it appears to be at the ental end of the atrium. One large prostate gland attached to the ental end of the atrial ampulla; a second prostate gland is not seen. Atrial ampulla sac-like (154–162 μm long, diameter 83–92 μm) and bent; atrial duct 43–46 μm long, diameter 36 μm. Atrial musculature very thin (2–3 μm), atrial epithelium 5–17 μm, thicker in the ectal section. Each atrium opens in a prominent round porophore (36 μm long, diameter 61 μm), where the penial chaeta is located ( Fig. 2E, G, H View FIGURE 2 ). Penis can be protruded from the porophore, length up to about 22 μm ( Fig. 2E View FIGURE 2 ).
One pair spermathecae with a very short, conical duct (27 μm long, 22 μm wide) and an elongate, globular ampulla (153–166 μm long, 106 μm maximum diameter), which contains a mass of sperm without forming discrete sperm bundles ( Fig. 2I View FIGURE 2 ).
Remarks. The new species is clearly ascribed to the genus Aberrantidrilus Martin, 2015 by the chaetal characteristics (simple pointed and bifid), position of the spermathecae in the postatrial segment, porophore and penis associated with a single, penial chaeta with the tip oriented forward. Our study material is limited, and the fact that an ectal prostate gland has not been seen in the new species is probably due to its small size, as for example in A. cuspis ( Erséus & Dumnicka, 1988) .
Prior to Martin et al. (2015), this new species would have been placed in Abyssidrilus , a genus erected by Erséus (1992) to include species of the subfamily Phallodrilinae with slender, simple-pointed and/or bifid chaetae, vas deferens entering the apical end of the atrium, and prostate glands attached separately at the most ental and most ectal ends of the atrium. However, Martin et al. (2015) erected the new genus Aberrantidrilus to accommodate all the freshwater subterranean species formerly classified as Abyssidrilus Erséus. In their review on the worldwide diversity of freshwater oligochaetes, Martin et al. (2008) reported 9 phallodriline genera, which included both marine and freshwater species. In this context, the validity of the genus Aberrantidrilus is stressed by the fact that the freshwater species formerly included in Abyssidrilus are clearly separated from the marine species (which remain in Abyssidrilus ) by the position of the spermathecal pores in the postatrial segment (XII), and by the presence of a single penial chaeta per bundle with the tip oriented forwards ( Martin et al. 2015). The marine Abyssidrilus species have typically 2 penial chaetae [except in A. stilus ( Erséus, 1986) with only one penial chaeta], which are oriented backwards, as indicated by Martin et al. (2015). Moreover, additional modified chaetae in spermathecal and prespermathecal segments (IX and X segments) are present in three out of nine Abyssidrilus species, but are absent in Aberrantidrilus . Finally, another interesting diagnostic and probably apomorphic character in Aberrantidrilus is the presence of ventral protuberances or porophores at the male pores, although, once more, the abyssal marine species Abyssidrilus stilus shares this character with the Aberrantidrilus species.
Compared with the previously described Aberrantidrilus species from subterranean freshwater habitats ( Table 2), the new species has the longest penial chaetae (3 times the mean length of somatic chaetae). Furthermore, the number of chaetae in posterior segments is higher in A. mihaljevici n. sp. than in congeners (5–7 vs up to 4). Total
length of the atrium appears to be within the range of other species described so far, but here the atrium has a greater diameter due to its sac-like shape, so its volume is larger than in congeners.
The new species is similar to A. cuspis in the shape of somatic chaetae, bifid anteriorly and simple-pointed posteriorly, but the spermathecal pores are located near septum 11/12 and in the longitudinal line of the ventral chaetae, whereas in A. cuspis they are located near the middle of the segment and laterally to the line of the ventral chaetae. Further comparison of A. mihaljevici n. sp. with A. cuspis is made difficult by apparent variability of characters and uncertain species boundaries of the latter. For example, the penial chaetae of A. cuspis are originally described as simple-pointed, whereas in Sambugar et al. (1999) they are bifid. These authors also report on a number of specimens with uncertain identity, identified as A. cf. cuspis , but with penial chaetae small or “may be absent”. The problematic variability of A. cuspis , which also probably includes material from Slovenia identified as “ Abyssidrilus sp. 1” by Giani et al. (2011), is presented and discussed in Martin et al. (2015), who hypothesize that A. cuspis may represent a complex of cryptic or “near-cryptic” species.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.