
Smithophis linearis, Vogel & Chen & Deepak & Gower & Shi & Ding & Hou, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4803.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:529351B7-252F-40B1-B03E-777A8B1A97BF |
|
persistent identifier |
https://treatment.plazi.org/id/D2181D7E-665B-4661-FF55-3325FB5EFDBA |
|
treatment provided by |
Plazi |
|
scientific name |
Smithophis linearis |
| status |
sp. nov. |
Smithophis linearis sp. nov.
( Figs. 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , 8–10 View FIGURE 8 View FIGURE 9 View FIGURE 10 )
Ablabes bicolor (Blyth, 1854) —Anderson (1879: 809).
Pseudocyclophis bicolor (Blyth, 1854) — Boulenger (1890: 300) (in part).
Rhabdops bicolor (Blyth, 1854) — Wall (1925: 810).
Rhabdops bicolor (Blyth, 1854) — Wall (1926: 561).
Rhabdops bicolor (Blyth, 1854) — Pope (1935: 176) (in part).
Rhabdops bicolor (Blyth, 1854) — Smith (1943: 328–329) (in part).
Rhabdops bicolor (Blyth, 1854) — Giri et al. (2019: 245) (in part).
Holotype. KIZ 059110 View Materials ( Figs. 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 ), an adult female, from Nabang Town (24°43'18.78"N, 97°35'03.12"E), Yingjiang County, Dehong Dai and Jingpo Autonomous Prefecture, Yunnan Province, People’s Republic of China ( Figs. 6 View FIGURE 6 , 7 View FIGURE 7 ), at an elevation of approximately 467 m a.s.l.; collected by Mian Hou on 10th August 2015. GoogleMaps
Paratypes (n = 3). ZSIK 4194 ( Fig. 8 View FIGURE 8 ), an adult male from Muangla (now Jiucheng Town: 24°44'29.12"N, 98°04'38.97"E, 849 m), Yunnan Province, People’s Republic of China, collected 1868 based on Anderson’s (1876) report of his expedition during which he visited Muangla in the first of two (1868 and 1875) expeditions. BMNH 1925.12.22.41 ( Fig. 9 View FIGURE 9 ) and BMNH 1925.12.22.42 ( Fig. 10 View FIGURE 10 ), adult males, from Huton [now Hutung], Kachin State, Myanmar (ca. 24.25° N, 97.52° E, ca. 1,500 m). Wall (1925, 1926) reported four specimens (as Rhabdops bicolor ) from Huton, Kachin Hills, Myanmar. Two specimens from Wall were accessioned into the BMNH collection on 22 December 1925, with locality data of “Huton, Kachin Hills”. On the basis of subcaudal scale counts, we assume that the two BMNH specimens are the two larger (of three) specimens reported by Wall (1926). We do not know the current whereabouts of the other two specimens reported by Wall (1925, 1926).
Diagnosis. Smithophis linearis sp. nov. differs from the two other nominal species of Smithophis in having six or more scales (excluding supralabials) contacting the eye (versus 5 in S. bicolor and 4 in S. atemporalis ) and in having a colour pattern comprising parallel, narrow, pale and very dark lines along the upper part of the body and tail. The new species is further distinguished from S. atemporalis by having (versus lacking) temporals. In terms of genetic divergence, Smithophis linearis sp. nov. differs from S. bicolor and S. atemporalis by a p-distance of>10% (cytb gene).
Etymology. The species epithet is the Latin adjective linearis (- e), meaning “with lines”, in reference to the narrow pale and dark lines along the upper part of the body.
Suggested common names. Jingpo mountain stream snake (English), Lined smithophis (English); Jingpo Gebirgsbachnatter (German). KỠḿẘ (Xiàn Wén Xī Shé) (Chinese).
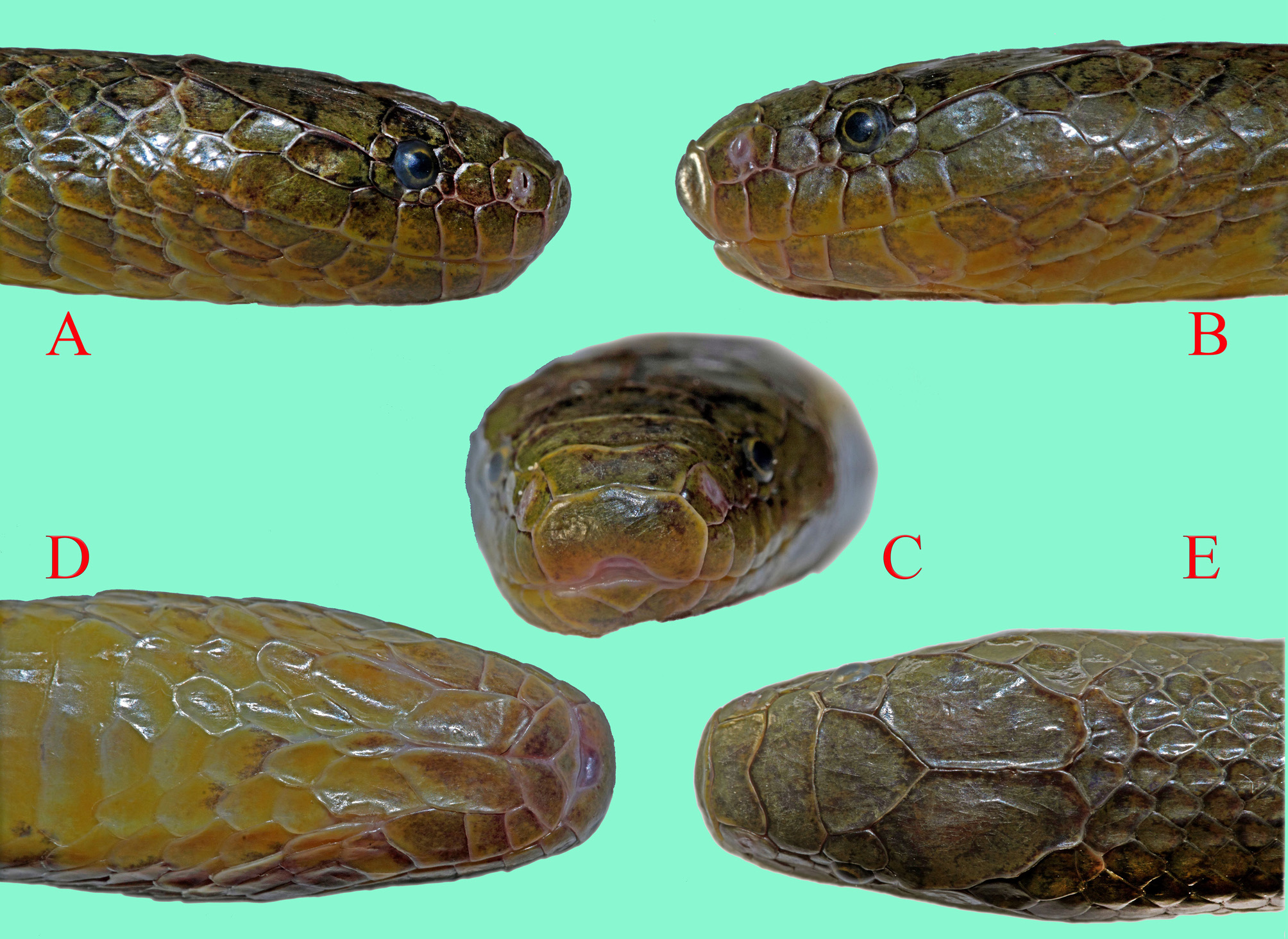
Description of holotype. Good condition. Body subcylindrical. Head short (3.3 % of SVL), barely wider than the thick anterior end of body, slightly distinct from neck; flattened anterior to eyes, dorsally covered with large scales; snout long, 31.1% of HL, 3.2 times longer than diameter of eye; snout blunt, subrectangular in dorsal view; nostrils anterolateral, crescentic and very narrow, almost reduced to a slit, strongly oblique, piercing the middle of nasal; eye small, its diameter about 0.7 times distance between eye and lip, pupil subcircular; tail long and gradually tapering.
Measurements. SVL: 493 mm; TaL: 116 mm; TL: 609 mm; ratio TaL/TL: 0.190; HL: 13.28 mm; SnL: 4.15 mm.
Body scalation. DSR: 17–17–16, relatively large, imbricate scales, smooth throughout the body. Dorsal scale rows undergo a single, simple reduction anteriorly:
3+4 (5)
19 ———–— 17
3+4 (5)
183 VEN (no preventrals); 59 SC, all paired; anal divided. ( Fig. 3 View FIGURE 3 ).
Head scalation. Rostral pentagonal, 1.5 times wider than high, visible from above; nasals large, distinctly directed forwards, subtriangular, much wider than high, obliquely divided by the long, narrow nostril and a short furrow below the nostril; internasal single, much broader than long, slightly tapering anteriorly; prefrontal single, much broader than long with anteromedial V-shaped projection, in contact with preocular and loreal on each side; frontal hexagonal with apex directed posteriorly, 1.5 times longer than prefrontal, 1.4 times wider than long; parietals long and wide, in contact for 1.1 times length of frontal; supraocular 1/1, small, about one quarter times as wide as frontal; loreal 1/1, subquadrangular, approximately 1.2 times higher than long, broadly in contact with internasal; preoculars 2/2; presubocular 1/1; suboculars 1/2; postsubocular 1/1; postocular 1/1; the second postsubocular largest of circumorbital scales after supraocular, subocular(s) smallest; supralabials 6/6; 1 st– 4 th higher than long, 6 th greatly elongate, much longer than 4 th and 5 th together; 1 st– 2 nd in contact with nasal, 2 nd– 3 rd contact loreal, 4 th SL contacts (post)subocular(s); temporals 1+1 / 1+1, anterior one largest and elongate, in broad contact with 5 th– 6 th SL and anterior part of parietal; infralabials 7/7, first pair in midline contact behind small mental, 1 st– 4 th ILs in contact with anterior genials, only 4 th IL in contact with posterior genials, 4 th IL distinctly the largest; posterior genials slightly shorter than anterior genials, not in contact with each other along midline ( Fig. 4 View FIGURE 4 ).
Coloration. In life, upper dorsal surface dark greyish-brown with bronze tints; lowest five dorsal scale rows (especially lowest two) paler bronze-brown or greenish-yellowish-brown. Upper and lower edges of each upper dorsal scale dark brown, forming 10 narrow dark lines, extending back from shortly behind head. Dark lines slightly wider and darker, more continuous dorsally; lines somewhat more conspicuous posteriorly and onto tail, extending to tail tip. On body, scales of 1 st DSR bronze-brown with substantial patches of the same pale ochre-brown colour as venter; smaller pale ochre-brown patches also on 2 nd DSR. Dorsal surface of tail mostly dark greyish-brown, as dorsum.
Head background colour mostly pale bronze-brown as sides of body, darker greyish-brown on snout and most of frontal. Supralabials dark greyish-brown with yellowish-ochre towards lower margins (only anteriorly so on 6 th SL), and posteroventral margins of first four SLs. Infralabials brownish with paler yellowish-ochre posterior and (to a lesser degree) upper margins. Except for infralabials, underside of head mostly pale, though anterior genials brownish centrally, anteriorly and medially. Some irregular brownish speckling
Venter of body uniform pale ochre, with diffuse brown speckles forming zig-zag along the upper margin of ventral scales. Ventral surface of tail pale ochre to cream, with some brown speckling, more so posteriorly, concentrated along midline in irregular zig-zag pattern along contacts between subcaudal scales of each side. In preservative, colour and pattern similar though somewhat darker, especially on venter.
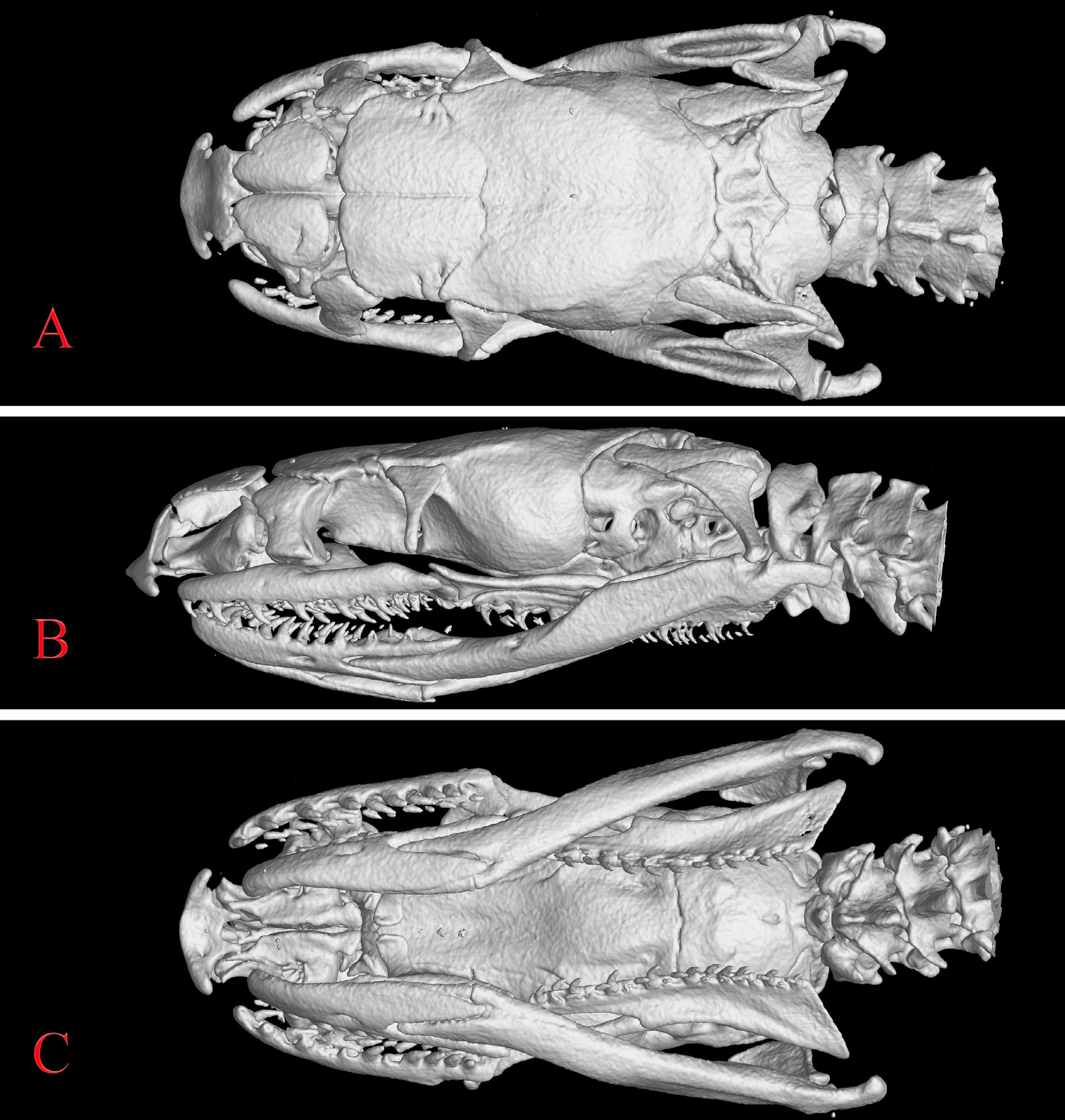
Dentition. Counts on each side estimated from microCT volume reconstruction ( Fig. 5 View FIGURE 5 ): 13 maxillary teeth, 5 th or 6 th largest, slightly smaller anteriorly and posteriorly; 8 palatine teeth; 16 pterygoid teeth; 17 dentary teeth, smaller posteriorly.
Variation among paratypes. Some meristic and metric data presented in Table 2. The paratypes closely resemble the holotype description except as noted here. BMNH 1925.12 .22.41 and BMNH 1925.12 .22.42 are in generally good condition, though both tend to dry quickly in air, and the former is slightly soft approximately three quarters of the distance between the snout tip and vent. BMNH 1925.12 .22.41 has two midventral incisions on the body, 47 mm (starting 220 mm behind the snout tip) and 23 mm long (ending 9 mm anterior to the vent), and one tail midventral incision 23 mm long (starting shortly behind the vent). BMNH 1925.12 .22.42 also has two body midventral incisions, 53 mm (starting 190 mm behind the snout tip) and 31 mm long (ending 7 mm anterior to the vent), and one tail midventral incision 16 mm long (starting shortly behind the vent). ZSIK 4194 lacks the tip of the tail and is dehydrated in parts, particularly towards the end of the tail and as indicated by a middorsal longitudinal groove on the posterior end of the body and anterior of the tail. In addition, two tags are tied tightly around and forming narrow constrictions of the anterior of the body, the posterior ends of the lower jaws are broken, the specimen is notably faded in patches, and several head shields lack their outer surface or have major artefactual creases .
DSR 17–17– 17 in both BMNH paratypes. Dorsal scale row reduction data not recorded for ZSIK 4194 ; both BMNH paratypes have a single reduction anteriorly from 19 to 17 DSR by VEN 2–3 with a merging of rows 1+2 (left) or 2+3 (right) ( BMNH 1925.12.22.41), and by VEN 4 with a merging of rows 3+4 (left) or 2+3 (right) ( BMNH 1925.12.22.42). BMNH 1925.12 .22.41 and ZSIK 4194 each have two preventrals (wide scales anterior to first ventral that do not contact first DSR on both sides) .
Eye diameter of the paratypes is greater relative to the distance between the eye and lip than in the holotype, though as in the holotype it is universally shorter than the eye-lip distance. In the BMNH paratypes the prefrontal is not entirely separate, being partially fused with the internasal on the right side of BMNH 1925.12 .22.41 and with the frontal on the left of BMNH 1925.12 .22.42. There is some variation in the circumorbital scalation. The three paratypes resemble the holotype in having a relatively large supraocular (though this is closer to one third the width of the frontal in the BMNH paratypes), in having one scale anterior and one scale posterior to the eye that lie below the supraocular and above the midline horizontal diameter through the eye. In addition to these three upper circumorbital scales, BMNH 1925.12 .22.41 and BMNH 1925.12 .22.42 have 4/4 and ZSIK 4194 has 3/3 other circumorbital scales. As in the holotype, the postsubocular (or ventral postocular, depending on preferred terminology) is the second largest circumorbital scale after the supraocular. BMNH 1925.12 .22.42 is unique among the type specimens in having an additional, small scale (that does not contact the eye or the lip) lying between SL2, SL3, loreal and lower circumorbital scales .
All paratypes differ from the holotype in the dimensions of the SLs, in that not all of SLs 1–4 are taller than long. In BMNH 1925.12 .22.41 only SL 1–3 are taller than long, in BMNH 1925.12 .22.42 only SL1, and in ZSIK 4194 only SL 1 and 3. The two BMNH paratypes have 5/5 (rather than 6/6) SLs, though all paratypes resemble the holotype in that the posteriormost SL is distinctly the longest. SL3 does not contact the loreal in BMNH 1925.12 .22.42. Infralabials 7/ 6 in BMNH 1925.12 .22.42. Temporals 1+2/1+ 1 in ZSIK 4194 . The posterior genials contact IL5 (as well as IL4 ) on the left of BMNH 1925.12 .22.41 and right of BMNH 1925.12 .22.42. The anterior genials contact only IL1–3 (not 1–4) on the left of BMNH 1925.12 .22.42. The posterior genials are similar in size to the anterior genials in BMNH 1925.12 .22.41 and 42 .
From microCT scan volume reconstructions,the tooth counts in BMNH 1925.12 . 22.41 and BMNH1925.12 . 22.42 are very similar for the maxilla (matching counts for the holotype) but variable for the dentary and especially the palate: 12–13 and 13 maxillary, 8 and 11 palatine, 14 and 16 pterygoid, and 14–15 and 17 dentary, respectively. Teeth not counted for ZSIK 4194 .
The colour is somewhat faded in all paratypes, though they all match the holotype in being pale ventrally and more intensely pigmented dorsally than laterally and ventrolaterally, and in having the distinct, lined pattern on the lateral and especially upper surface. The darker pigment on the underside of the anterior of the head does not extend onto the posterior genials in BMNH 1925.12 .22.41.
Remarks. In addition to the generic differences between Smithophis and Opisthotropis , S. linearis sp. nov. differs from all species of Opisthotropis as summarised in Appendix 4. teynieì et al. (2013) provide an identification key to the species of Opisthotropis , together with Parahelicops annamensis (now Hebius annamensis ) ( Kizirian et al. 2018), Paratapinophis praemaxillaris and “ Opisthotropis ” boonsongi (now Isanophis boonsongi ) ( David et al. 2015). using this key, Smithophis linearis sp. nov. would ‘key out’ as (most similar to) O. maxwelli in couplet 14, though these two species differ clearly in colour pattern (lined in S. linearis sp. nov., uniform in O. maxwelli ), and in number of ventrals (183–191 in S. linearis sp. nov., 147–155 in O. maxwelli ). Using Ren et al. ’s (2017) more recent key, S. linearis sp. nov. does not key out or would lead to O. lateralis . Smithophis linearis sp. nov. differs clearly from O. lateralis in having a lined dorsal pattern and substantially fewer supralabials (6 vs. 9 or more). If Ziegler et al. ’s (2017) updated description of O. tamdaoensis (anterior temporal scales 1–2 instead of only 2) is added to Ren et al. ’s (2017) key, S. linearis sp. nov. would key out as O. tamdaoensis . Smithophis linearis sp. nov. differs clearly from O. tamdaoensis in having more subcaudals (59–64 vs. 48–51), fewer temporals (1+1 vs. 1–2+2–4), fewer supralabials (6 vs. 9, rarely 8) and strongly lined colouration vs. uniformly dark with a black lateral stripe
Distribution. Smithophis linearis sp. nov. is potentially distributed in an area with a radius of at least ca. 40 km, based on the three known localities. The type locality is a lower hilly region located in Nabang Town, Yingjiang County, Dehong Dai and Jingpo Autonomous Prefecture, in Southwest Yunnan Province, China, very close to the border with Myanmar. The paratypes are from Jiucheng Town, Yunnan Province, People’s Republic of China and Huton [now Hutung], Kachin State, Myanmar.
Natural history and conservation. The holotype of Smithophis linearis sp. nov. was found in tropical evergreen forest at an elevation of about 467 m a.s.l. The specimen was found at night on a rock along a small rocky stream. Wall (1926) recorded that among his specimens from Myanmar, one had consumed “what appeared to be a slug” and another an “arachnoid” .
The climate of the area of the type locality is influenced by the India Ocean monsoon with four seasons. Annual precipitation is approximately 1,500 mm, annual average sunshine is 2,350 h, mean annual temperature is 18–20 °C, with maximum temperature up to 36.8 °C ( Wei, 2011; Yang and Du, 2006; Yin, 2003). The fauna of this region belongs to the western Gaoligong Mountain subregion in the region of the East Himalayan-Gaoligong Mountains. This region is characterized by species inhabiting the Brahmaputra and upper Ayeyarwady (Irrawaddy) River basins. The western Gaoligong Mountain subregion is part of the upper Ayeyarwady River basin, the central-south range of the Gaoligong Mountains that was usually called “Kakhyen Hills” [= Kachin Hills] by western explorers in the 1800s (e.g. Anderson 1876).
More than 103 species of mammals, 383 species of birds and 98 species of amphibians and reptiles have been recorded from this area ( Fan et al. 2017; Hu & Han 2007; Hou et al. 2017; Qu & Yang 1996, 1998; Wei 2011; Yang et al. 2008; Yang & Rao 1996; Yang & Du 2006; Yin 2003; Zhao1998, 2006). Characteristic reptiles include Calotes mystaceus Duméril & Bibron, 1837 , C. jerdoni Günther, 1870 , Japa lura variegata Gray, 1853, Pseudocalotes kakhienensis (Anderson, 1879) , Varanus bengalensis irrawadicus Yang & Li, 1987 , Atretium yunnanensis Anderson, 1879 , Dendrelaphis subocularis ( Boulenger, 1888) , Lycodon fasciatus (Anderson, 1879) and Oligodon albocinctus (Cantor, 1839) .
The Tongbiguan Nature Reserve (23°54′- 24°51′ N, 97°31′- 98°05′ E: Wei, 2011; Yang and Du, 2006; Yin, 2003) lies close to the known distribution of Smithophis linearis sp. nov. so the new species potentially extends into this protected area. However, S. linearis sp. nov. has not been recorded in two of its localities for 95–151 years, so confirming its presence in these sites and obtaining better data on distribution and possible threats will likely be required to formulate an IUCN Red List conservation assessment of other than Data Deficient.







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Smithophis linearis
| Vogel, Gernot, Chen, Zening, Deepak, V., Gower, David J., Shi, Jingsong, Ding, Li & Hou, Mian 2020 |
Rhabdops bicolor (Blyth, 1854)
| Giri, V. B. & Gower, D. J. & Das, A. & Lalremsanga, H. T. & Lalronunga, S. & Captain, A. & Deepak, V. 2019: 245 |
Rhabdops bicolor (Blyth, 1854)
| Smith, M. A. 1943: 328 |
Rhabdops bicolor (Blyth, 1854)
| Pope, C. H. 1935: 176 |
Rhabdops bicolor (Blyth, 1854)
| Wall, F. 1926: 561 |
Rhabdops bicolor (Blyth, 1854)
| Wall, F. 1925: 810 |