
Neocallichirus Sakai, 1988
|
publication ID |
https://doi.org/10.5281/zenodo.209484 |
|
DOI |
https://doi.org/10.5281/zenodo.5618124 |
|
persistent identifier |
https://treatment.plazi.org/id/D03A87F0-FFB6-FFDC-FF5E-83F21C256E64 |
|
treatment provided by |
Plazi (2016-04-12 07:35:44, last updated 2024-11-29 17:05:18) |
|
scientific name |
Neocallichirus Sakai, 1988 |
| status |
|
Genus Neocallichirus Sakai, 1988
Type species. Neocallichirus horneri Sakai, 1988 by original designation.
Remarks. The genus was erected by Sakai (1988) to accomodate callianassid forms characterized by the following features: carapace with well-defined dorsal oval, but without rostral carina and cardiac prominence; 3rd maxilliped without exopod, endopod subpediform, propodus expanded, over 3 times broader than dactylus; uropodal endopod broadened posteriorly; telson about as long as broad, about as long as uropods. These characters are, however, of very poor fossilization potential, and as Sakai (1988) stated, the form of the 3rd maxilliped alone is not specific to Neocallichirus , but is similar in several other genera, and therefore always a combination of characters has to be used for generic assignment. Later, Manning & Felder (1991) reconsidered the diagnosis of Neocallichirus and added also the characters on the chelipeds, which are common in the fossil record and therefore usable for palaeontologists too. They argued for the taxonomic importance of the shape of the merus (e.g. presence/absence of meral hook), which is significant for many callianassid genera. Schweitzer & Feldmann (2002) and Schweitzer et al. (2006 a) discussed usage of the characters on the major cheliped in the fossil record for assignment to the genus Neocallichirus . They noted that the carpus is variable in shape; however, it is not rectangular and typically has a rounded lower margin. The propodus has often serrated distal margin. The fixed finger tends to be edentulous, whereas the dactylus is stout and heavily armed with a triangular or rectangular tooth near the proximal end of the element.
The merus of members of Neocallichirus is variable in shape but is always serrate along the lower margin. According to Manning & Felder (1991), it lacks a meral spine or hook; according to Sakai (1999, 2005) it is with or without meral hook (mainly due to the synonymisation with the genus Sergio Manning & Lemaitre, 1994 ). Sakai (2011) stated that the merus is with or without ventral convexity.
There are several genera different from Neocallichirus , namely Sergio , Podocallichirus Sakai, 1999 , and Grynaminna Poore, 2000 , which in general share characters on the chelipeds mentioned above. It is typical that such a morphology, i.e. generalized cheliped morphology embracing all above mentioned genera, is in palaeontological literature connected virtually with Neocallichirus only. This can be documented by currently recognized fossil taxa assigned to respective genera. According to De Grave et al. (2009) and Schweitzer et al. (2010) there are 18 fossil species of Neocallichirus , but only one fossil species of Podocallichirus (known from the Pleistocene strata of Japan; see Karasawa et al. 2006) and no fossil Grynaminna (see Hyžný & Karasawa in press for a different view). One extant species of Sergio is also known from the fossil state ( Portell & Agnew 2004).
Much confusion has been created by Sakai (1999, 2005) when he synonymized Sergio with Neocallichirus and Grynaminna with Podocallichirus (see also Sakai 2011 for a different view on the taxonomy of these genera). Thus, we can talk about several different taxonomic concepts of Neocallichirus presented by Sakai (1988, 1999, 2005, 2011) and Manning & Felder (1991). Identifying the fossil material as a member of any of the above mentioned genera strongly depends on taxonomy adopted. Therefore it is possible that the genus Neocallichirus as usually recognized in the fossil record is a mixture of closely allied genera. The issue was recently discussed by Hyžný & Karasawa (in press).
Neocallichirus brocchii (L ő renthey, 1897) new combination ( Figs 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
Calianassa Brocchii Lőrenthey, 1897: 161 , 168, 169; Lőrenthey, 1898a: 106, 114, 115; Lőrenthey, 1898b: 132 –134, 155, pl. 9, fig. 5; Lőrenthey, 1898b: 104 –105, pl. 9, fig. 5; Lőrenthey in Lőrenthey & Beurlen, 1929: 64, pl. 2, fig. 13;
Callianassa Brocchii. View in CoL — Glaessner, 1929: 77.
? Calianassa cf. Brocchii . — Lőrenthey in Lőrenthey & Beurlen, 1929: 69.
Callianassa brocchii. View in CoL — Müller, 1979: 274; Schweitzer et al., 2010: 34.
‘ Callianassa View in CoL ’ brocchii . — Müller, 1984: 51, pl. 3 fig.3.
" Callianassa View in CoL " brocchi (sic). — Hyžný, 2011a: 41, Table 2 View TABLE 2 .; Hyžný, 2011b: 167, Table 1 View TABLE 1 .
Emended diagnosis. Strongly heterochelous callichirine ghost shrimp with chelipeds without pronounced tuberculation. Merus of major cheliped ovoid with lower margin possesssing a toothed blade consisting of up to 12 teeth; carpus broad with rounded and serrated lower margin; propodus tapering distally with serrated lower margin, the palm subequal in length and height, distal margin rounded and serrated, occlusal margin of fixed finger armed with small teeth; dactylus long and slender with occlusal margin forming a well developed keel with long, quadrate, blunt tooth proximally and one indented, sharp tooth distally followed with several tiny teeth, tip forming sharp hook. Carpus of minor cheliped slightly higher than propodus; propodus slender, tapering distally, fixed finger about as long as manus; fingers without armature, occlusal margin of dactylus forming weak serrated keel.
Emended description. Major cheliped massive, located on either right or left side of body, smooth, without pronounced tuberculation on lateral surfaces; outer surface usually vaulted. Two different morphotypes were identified in the nature of major cheliped; robust morphotype is relatively larger than slender morphotype. Ischium is slender, longer than high, tapering proximally, not well preserved.
Major chela robust morphotype: Merus ovoid in shape, longer than high (L/H = 1.5–1.7), highest proximally; lower margin with toothed blade (up to 12 teeth present); upper margin slightly convex ( Figs 2 View FIGURE 2 G, 4F–H, J, K). Carpus broad, massive, approximately as long as high (L/H = 0.8–1.1); upper margin straight; lower margin rounded forming one edge with proximal margin ( Figs 4 View FIGURE 4 A–E), serrated with tiny denticles ( Fig. 4 View FIGURE 4 D); upper and lower margin forming prominent keel curved inward. Propodus massive, longer than high, slightly longer than carpus, tapering distally ( Figs 2 View FIGURE 2 A, 3); palm of the propodus subequal in length and height; upper margin slightly convex; lower margin straight, serrated ( Figs 3 View FIGURE 3 A, O); upper and lower margins keeled, keel missing on fixed finger; distal margin rounded and serrated with up to 10 small denticles on each side of articulation with dactylus, serrated incision present just below the articulation with dactylus ( Figs 2 View FIGURE 2 A, 3D, J, M). Fixed finger slender, triangular in shape; occlusal margin armed with small teeth, forming serrated keel on the outer side of fixed finger from distal view ( Figs 2 View FIGURE 2 E, 3S); tip curved slightly upward; lower margin serrated. Dactylus long and slender, occlusal margin forming sharp keel with long quadrate blunt tooth proximally and one indented sharp tooth distally followed with several tiny teeth ( Figs 2 View FIGURE 2 F, 3E–I); tip bent slightly downward or forming sharp hook ( Fig. 3 View FIGURE 3 G).
Major chela slender morphotype: Merus ovoid in shape, longer than high, highest at midlength; lower margin finely serrated without distinct tooth blade ( Figs 2 View FIGURE 2 H, 4I). Carpus higher than long (L/H = 0.70–0.85) ( Figs 2 View FIGURE 2 B, 3P, 6C), keels on upper and lower margins not as pronounced as in robust morphotype; lower margin serrated. Propodus distinctly longer than carpus, general shape much the same as in robust morphotype ( Figs 2 View FIGURE 2 B, 3B, 6C). Fixed finger with serrated keel on occlusal margin positioned in the middle ( Figs 2 View FIGURE 2 D, 3Q, R) instead of lateral position of robust morphotype. Dactylus similarly shaped as in robust morphotype ( Fig. 3 View FIGURE 3 B).
Minor cheliped more slender than major one, smooth ( Fig. 2 View FIGURE 2 C). Ischium and merus not well preserved. Carpus slightly longer than high, higher than propodus ( Figs 2 View FIGURE 2 C, 6B); upper and lower margins convex. The palm of the propodus as long as high; distal margin without serration ( Figs 5 View FIGURE 5 A–D); lower margin of propodus slightly convex at the point where fixed finger beggins. Fixed finger about as long as palm, slender, forming sharp tip, occlusal margin not armed. Dactylus long, slender; occlusal margin forming faint finely serrated keel, no bigger teeth present ( Fig. 5 View FIGURE 5 E).
Variations. The studied material consists of the two morphotypes described above. The overall shape of the propodus and morphology of the dactylus and minor chela clearly demonstrate that both morphs belong to the same species. Both morphotypes occur together in the same strata, so they represent the same population (or sequence of populations). The recognized morphs may mirror sexual dimorphism (slender morphotype being a female one); however, it is very difficult to determine sex in the fossil state, especially when dealing with Neocallichirus . No obvious sexual dimorphism has been observed in several extant species (see Dworschak 2011a, b). Thus, the interpretation of morphs as sexual dimorphism should be considered as a preliminary one. Possible attribution of the slender morphotype to female individuals is based mostly on other callianassid taxa in which mature males usually possess more robust chelipeds than females (e.g., Felder & Lovett 1989 and references therein).
The merus shows variations in the number of teeth on its lower margin which seems to be a consequence of growth (more teeth are present on larger meri) rather than real variability. The variation in meral shape, notably the development of the meral blade, can be documented in several extant Neocallichirus species, e.g. in N. grandimanus ( Gibbes, 1850) (compare e.g. Manning 1987: fig. 2; Lemaitre & Ramos 1992: fig. 5; Blanco-Rambla 2000: fig. 2) and N. jousseaumei ( Nobili, 1904) (compare e.g. Nobili 1906: fig. 2; Kensley 1976: fig. 2; Dworschak 2011 a: figs. 1–4). In this respect the merus of the slender morphotype of Neocallichirus brocchii comb. nov., without a pronounced meral blade ( Fig. 2 View FIGURE 2 H), is similar to the merus of N. grandimanus depicted by Lemaitre & Ramos (1992: fig. 5d), whereas the merus of the robust morphotype with a well developed meral blade ( Fig. 2 View FIGURE 2 G) can be compared with the merus of N. grandimanus depicted by Blanco-Rambla (2000: fig. 2c).
There are slight variations in the outline of the distal margin of the propodus. Variations exist mainly in the nature of the incision under dactylus articulation. In the robust morphotype the incision is usually deeper and more rounded (e.g. Figs 3 View FIGURE 3 D, J, M), whereas in the slender morphotype, it is sometimes completely missing.
Other variations concern the cutting edge of the dactylus. The proximal quadrate tooth is usually divided in two secondary blunt teeth, the distal one being broader. The subdivision is, however, not always distinct and also the shape of the subdivided teeth is variable ( Fig. 2 View FIGURE 2 F).
No obvious allometric growth in the nature of any cheliped element has been recognized, although there is one isolated propodus in the present collection (KGP-MH DH066; Fig. 3 View FIGURE 3 T) which shows a propodus length/height ratio (L/H = 0.6) rather different from all other studied specimens. The nature of the fixed finger (position of serrated keel) resembles that of the slender morphotype. The specimen itself is, however, much bigger than the usual size of the slender morphotype propodus.
Material examined. FI M.2329 (holotype – left propodus of robust morphotype articulated with dactylus); KGP-MH DH-001–KGP-MH DH-015; M.86.249 (1 left and 1 right fragmented propodi; SNM Z- 7943 (collective number), SNM Z- 16419 (collective number), SNM Z- 16433 (collective number), SNM Z- 37532– SNM Z- 37553; PCMH-001–PCMH-003; PCRB DH-001– PCRB DH-016. For measurements and all the details see Table 1 View TABLE 1 .
merus carpus propodus dactylus SNM Z- 37548 unknown R 8.7 5.0 - - - - - continued next page Occurrence. The species has been reported only from the Serravallian (Middle Miocene), specifically from the upper 'Badenian' of the Rákos Formation – localities Rákos, Budafok and Gyakorló út in the Budapest City area ( Hungary) ( Lőrenthey & Beurlen 1929; Müller 1984); and the lower 'Sarmatian' of the Studienka Formation – locality Dúbravská hlavica in the Bratislava district ( Slovakia)
( Hyžný 2011 a, b; this contribution).
Remarks. The species possesses a combination of characters which is quite typical for Neocallichirus : serrated lower margins of the merus, carpus and propodus; rounded lower margin of the carpus; upper margin of the propodus converging distally; an edentulous fixed finger (which is only finely denticulated), and an armed dactylus with hooked tip.
In his major work, Müller (1984: 51) already noted the morphological similarity of Callianassa brocchii to C. natalensis Barnard, 1947 . Müller (1984) examined only the holotype of C. brocchii and five additional propodi from the Budapest City localities (see above). At that time no carpi or meri were known, or at least were not attributed to C. brocchii .
Müller (1984: 51) stated that Callianassa brocchii can be differentiated from similar extant species ( Callianassa gilchristi Barnard, 1947 , C. natalensis ) on the basis of "the saw-like lower edge". This is not correct, however, as C. natalensis does possess serration on the lower margin of the propodus (see e.g. Dworschak 2011 a: fig. 5D, E) as do Neocallichirus species.
Neocallichirus brocchii comb. nov. is morphologically very similar to the extant Neocallichirus lemaitrei Manning, 1993 , and allied species (see also comparison with N. grandimanus and N. jousseaumei above). The armature of the dactylus is similar to extant N. cacahuate Felder & Manning, 1995 , N. jousseaumei , N. lemaitrei , N. monodi (de Saint Laurent & Le Loeuff, 1979) and N. natalensis . In fossil taxa the variation in the shape of the dactylus is not very well known; however, none of those described can be fully compared to Neocallichirus brocchii comb. nov. The combination of characters as presented in the diagnosis and description is unique for this species, although some similarities can be found with Neocallichirus aetodes Schweitzer, Iturralde-Vinent, Hetler & Velez- Juarbe, 2006 from the Lower Oligocene of Puerto Rico and N. hattai Karasawa & Nakagawa, 2010 from the Lower to Middle Miocene of Japan.
The original description of Callianassa brocchii is based on a single specimen of left major propodus articulated with dactylus; Lőrenthey (1898 a) and Lőrenthey & Beurlen (1929) erroneously treated it as a right one, although the accompanying figures clearly show it as being left, as re-examination of the type material also confirmed. Lőrenthey in Lőrenthey & Beurlen (1929: 65) mentioned another two specimens, which may probably be assigned to this species, although no figures of them were added. Another specimen was reported by Beurlen in Lőrenthey & Beurlen (1929: 69). He classified the material as Callianassa cf. brocchii , however, he stated that it morphologically conforms with the description of C. brocchii . As no figure of the material was added, we are hesitant to assign the material to the species with certainty. Interestingly in the description it is mentioned that "Form des Carpus, die Grösse, der Index, soweit es das abgebrochene Stück erkennen lässt, stimmen volkommen mit dieser Art überein" ( As far as the broken specimen can be identified, the form of carpus, its size, and the fixed finger correspond precisely to this species). As C. brocchii has been described on the basis of an articulated propodus and dactylus only, and no carpus was described in the original description, we assume that Beurlen made a mistake and meant, in fact, propodus instead of carpus.
There is much confusion in citing Lőrenthey's works. As stated by Müller (1984: 32): "Lőrenthey's first papers were published in parallel Hungarian and German versions with identical contents. He presented the same data in preliminary publications without figures ( Lőrenthey 1897, 1898a), and in definite papers with excellent figures and detailed descriptions ( Lőrenthey 1898b, c)". Callianassa brocchii was first described as a new species in such a preliminary publication in 1897. In this paper a short description with comparison with similar taxa can be found which satisfies the ICZN rules for a new species designation. Later the description of C. brocchii was refined and figures were added ( Lőrenthey 1898b, c). Thus, the year 1897 is considered as the year of description of Callianassa brocchii contrary to Glaessner (1929) and Schweitzer et al. (2010) where the species appeared as described in 1898 (see also discussion in Müller 1984: 32).
Müller (1984: 50) erroneously assumed that the handedness is quite typical for every callianassid species: "Most of recent Callianassids (sic!) are strongly heterochelous. Surprisingly, in the diverse 'Badenian' material no evidence of heterochely was observed, as right and left chelae are found in same proportion and in same size as well, with the exception of C. brocchii ". It should be noted here that Müller (1984) examined only six specimens of C. brocchii comparing to often tens and hundreds of specimens in other " Callianassa " species. Therefore, it seemed to him that C. brocchii was the only heterochelous species among 'Badenian' callianassid forms.
Schweitzer, C. E., Gonzalez-Barba, G., Feldmann, R. M. & Waugh, D. A. (2006 a) Decapoda (Thalassinidea and Paguroidea) from the Eocene Bateque and Tepetate Formations, Baja California Sur, Mexico: systematics, cuticle microstructure, and paleoecology. Annals of Carnegie Museum, 74 (4), 275 - 293.
De Grave, S., Pentcheff, N. D., Ahyong, S. T., Chan, T. - Y., Crandall, K. A., Dworschak, P. C., Felder, D. L., Feldmann, R. M., Fransen, C. H. J. M., Goulding, L. Y. D., Lemaitre, R., Low, M. E. Y., Martin, J. W., Ng, P. K. L., Schweitzer, C. E., Tan, S. H., Tshudy, D. & Wetzer, R. (2009) A classification of living and fossil genera of decapod crustaceans. The Raffles Bulletin of Zoology Supplements, 21, 1 - 109.
Schweitzer, C. E., Feldmann, R. M., Garassino, A., Karasawa, H. & Schweigert, G. (2010) Systematic list of fossil decapod crustacean species. Crustaceana Monographs, 10, 1 - 222.
Barnard, K. H. (1947) Descriptions of new species of South African decapod Crustacea, with notes on synonymy and new records. Annals and Magazine of Natural History, series 11, 13, 361 - 392. [Dated 1946, published 1947]
Blanco-Rambla, J. P. (2000) Cuatro nuevos registros de especies del genero Neocallichirus (Crustacea: Decapoda: Callianassidae) para aguas venezolanas. Boletin del Instituto Oceanographico de Venezuela, Universidad de Oriente 39 (1 - 2), 71 - 78.
Dworschak, P. C. (2011 a) Redescription of Callianassa jousseaumei Nobili, 1904, a junior (sic!) subjective synonym of Callianassa indica de Man, 1905 with description of a new species of Neocallichirus (Decapoda: Axiidea: Callianassidae). Zootaxa, 2746, 1 - 19.
Dworschak, P. C. (2011 b) Redescription of Callianassa vigilax de Man, 1916, a subjective synonym of Neocallichirus denticulatus Ngoc-Ho, 1994 (Crustacea: Decapoda: Callianassidae). Annalen des Naturhistorischen Museums in Wien, B 112, 137 - 151.
Felder, D. L. & Lovett, D. L. (1989) Relative growth and sexual maturation in the estuarine ghost shrimp Callianassa louisianensis Schmitt, 1935. Journal of Crustacean Biology, 9 (4), 540 - 553.
Felder, D. L. & Manning, R. B. (1995) Neocallichirus cacahuate, a new species of ghost shrimp from the Atlantic coast of Florida, with reexamination of N. grandimana and N. lemaitrei (Crustacea: Decapoda: Callianassidae). Proceedings of the Biological Society of Washington, 108 (3), 477 - 490.
Gibbes, L. R. (1850) On the carcinological collections of the United States, and an enumeration of the species contained in them, with notes on the most remarkable, and descriptions of new species. Proceedings of the American Association for the Advancement of Science, 3 rd meeting, 167 - 201.
Glaessner, M. F. (1929) Crustacea Decapoda. In. Pompeckj, F. J. (Ed.), Fossilium catalogus, 1: Animalium, 41, 1 - 464. W. Junk, Berlin.
Hyzny, M. (2011 a) In situ mud shrimps (Decapoda: Axiidea: Callianassidae) preserved within their burrows from the middle Miocene of the Central Paratethys. Bulletin of the Mizunami Fossil Museum, 37, 37 - 46.
Hyzny M. (2011 b) Synopsis of fossil decapod crustaceans from Slovakia (Western Carpathians). Neues Jahrbuch fur Geologie und Palaontologie, Abhandlungen, 260 (2), 165 - 171.
Karasawa, H., Tanaka, T., Kobayashi, N., Goda, T., Ohira, N. & Shinya, J. (2006) Podocallichirus grandis (Crustacea: Decapoda: Thalassinidea) preserved within burrows from the middle Pleistocene Atsumi Group of Aichi Prefecture, Japan. Bulletin of the Mizunami Fossil Museum, 33, 127 - 133. [In Japanese]
Karasawa, H. & Nakagawa, T. (2010) A new ghost shrimp (Decapoda: Thalassinidea) from the Miocene Kunimi Formation, Fukui Prefecture, Japan. Bulletin of the Mizunami Fossil Museum, 36, 31 - 36.
Kensley, B. (1976) Records of mud-prawns (genus Callianassa) from South Africa and Mauritius (Crustacea, Decapoda, Thalassinidea). Annals of the South African Museum, 69, 47 - 58.
Lemaitre, R. & Ramos, G. E. (1992) A collectio of Thalassinidea (Crustacea: Decapoda) from the Pacific coast of Colombia, with description of a new species and a checklist of Eastern Pacific species. Proceedings of the Biological Society of Washington, 105, 343 - 358.
Lorenthey, E. (1897) Adatok Magyarorszag harmadkoru rak faunajahoz. Mathematikai es Termeszettudomanyi Ertesito, 15, 149 - 169.
Lorenthey, E. (1898 a) Beitrage zur Decapodenfauna des ungarischen Tertiars. Mathematischen und Naturwissenschaftlichen Berichte aus Ungarn, 14, 92 - 115.
Lorenthey, E. (1898 b) Adatok Magyarorszag harmadkoru rak-faunajahoz. Mathematikai es Termeszettudomanyi Kozllemenyek, 27 (2), 103 - 271.
Lorenthey, E. (1898 c) Beitrage zur Decapodenfauna des Ungarischen Tertiars. Termeszetrajzi Fuzetek, 21, 1 - 133.
Lorenthey, E. & Beurlen, K. (1929) Die fossilen Dekapoden der Lander der Ungarischen Krone. Geologica Hungarica, Series Palaeontologica, 3, 1 - 421.
Manning, R. B. (1987) Notes on western Atlantic Callianassidae (Crustacea: Decapoda: Thalassinidea). Proceedings of the Biological Society of Washington, 100 (2), 386 - 401.
Manning, R. B. & Felder, D. L. (1991) Revision of the American Callianassidae (Crustacea: Decapoda: Thalassinidea). Proceedings of the Biological Society of Washington, 104 (4), 764 - 792.
Manning, R. B. (1993) Two new species of Neocallichirus from the Caribbean Sea (Crustacea: Decapoda: Callianassidae). Proceedings of the Biological Society of Washington, 106, 106 - 114.
Manning, R. B. & Lemaitre, R. (1994) Sergio, a new genus of ghost shrimp from the Americas (Crustacea: Decapoda: Callianassidae). Nauplius, 1, 39 - 43.
Muller, P. (1979) Decapoda (Crustacea) fauna a budapesti miocenbol (5). Foldtani Kozlony, Bulletin of the Hungarian Geological Society, 108, 272 - 318.
Muller, P. (1984) Decapod Crustacea of the Badenian. Geologica Hungarica, Series Palaeontologica, 42, 3 - 317.
Nobili, G. (1904) Diagnoses preliminaires de vingt-huit especes nouvelles de Stomatopodes et Decapodes Macroures de la Mer Rouge. Bulletin du Museum d'Histoire Naturelle de Paris, 10, 230 - 238.
Nobili, G. (1906) Faune carcinologique de la Mer Rouge: Decapodes et Stomatopodes. Annales des Sciences Naturelles, Zoologie, (9) 4, 1 - 347.
Poore, G. C. B. (2000) A new genus and species of callianassid ghost shrimp from Kyushu, Japan (Decapoda: Thalassinidea).
Portell, R. W. & Agnew, J. G. (2004) Pliocene and Pleistocene decapod crustaceans. Florida Fossil Invertebrates, 4, 1 - 29.
Saint Laurent, M. de & Le Loeuff, P. (1979) Upogebiidae et Callianassidae. Crustaces Decapodes Thalassinidea, 1. Campagnes de la Calypso au large des cotes Atlantiques africaines (1956 et 1959) (suite), 22. Resultats scientifiques des campagnes de la Calypso, 11, 29 - 101.
Sakai, K. (1988) A new genus and five new species of Callianassidae (Crustacea: Decapoda: Thalassinidea) from Northern Australia. The Beagle, Records of the Northern Territory Museum of Arts and Sciences, 5 (1), 51 - 69.
Sakai, K. (1999) Synopsis of the family Callianassidae, with keys to subfamilies, genera and species, and the description of new taxa (Crustacea: Decapoda: Thalassinidea). Zoologische Verhandelingen (Leiden), 326, 1 - 152.
Sakai, K. (2005) Callianassoidea of the world (Decapoda: Thalassinidea). Crustaceana Monographs, 4, 1 - 285.
Sakai, K. (2011) Axioidea of the World and a Reconsideration of the Callianassoidea (Decapoda, Thalassinidea, Callianassida). Crustaceana Monographs, 13, 1 - 520.
Schweitzer, C. E. & Feldmann, R. M. (2002) New Eocene decapods (Thalassinidea and Brachyura) from Southern California. Journal of Crustacean Biology, 22 (4), 938 - 967.
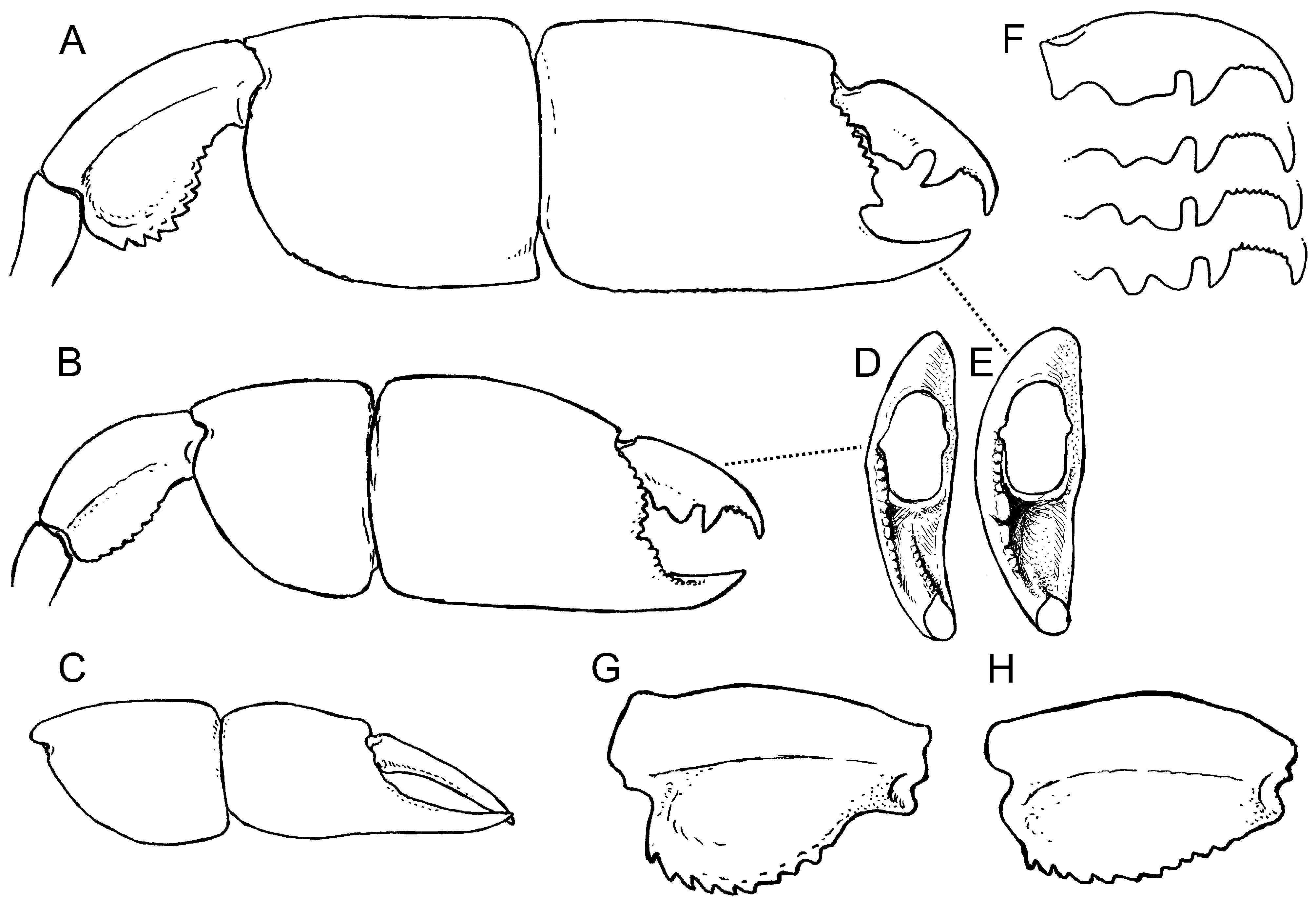
FIGURE 2. Neocallichirus brocchii (Lőrenthey, 1897) new combination, reconstructions: A, major cheliped of robust morphotype (outer view); B, major cheliped of slender morphotype (outer view); C, minor cheliped (outer view); D – E, distal view of propodus of slender (D) and robust (E) morphotype, note differences in the development of medial keel on the fixed finger; dactyli and fixed finger tips are not depicted; F, variation in the armature of fixed finger; G – H, outer view on merus of robust (G) and slender (H) morphotype. Reconstructions are based on several different specimens; figures A – F are in scale.
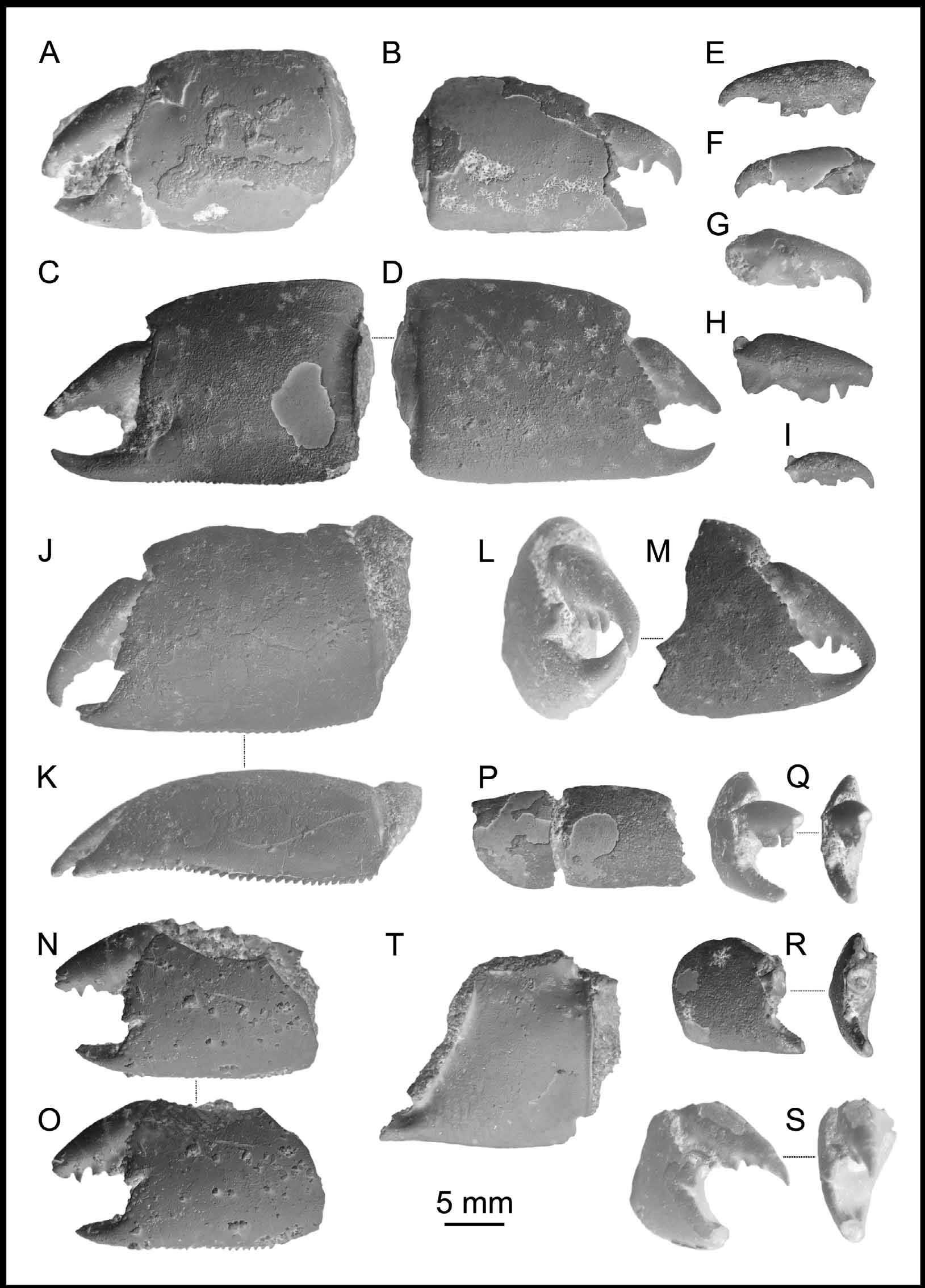
FIGURE 3. Neocallichirus brocchii (Lőrenthey, 1897) new combination, major chelae (propodi and dactyli): A, left chela (robust morphotype), KGP-MH DH 001; B, right chela (slender morphotype), PCMH- 001; C – D, right chela from inner and outer view (robust morphotype), PCRB-DH 008; E, left dactylus, PCRB-DH 001; F, left dactylus, PCRB-DH 002; G, right dactylus, KGP-MH DH 012; H, right dactylus, PCRB-DH 004; I, right dactylus, PCRB-DH 003; J, left chela (robust morphotype), KGP-MH DH 064, note serration on the lower margin (K); L – M, fragmentary right chela (robust morphotype), PCRB-DH 005; N, left chela (robust morphotype), SNM Z- 37534, note serration on the lower margin (O); P, right chela (slender morphotype), PCRB-DH 010, note short carpus; Q, fragmented right chela (slender morphotype), PCMH- 003, note occlusal keel on the fixed finger positioned medially; R, right chela (slender morphotype), PCRB-DH 007, note occlusal keel on the fixed finger positioned medially; S, fragmented right chela (robust morphotype), PCMH- 002, note occlusal keel on the fixed finger positioned laterally; T, right chela, KGP-MH DH- 66, note the unusual length / height ratio. All specimens come from the Dúbravská hlavica locality (Vienna Basin, Slovakia), Middle Miocene. All specimens are to scale and were covered with ammonium chloride prior to photography.

FIGURE 4. Neocallichirus brocchii (Lőrenthey, 1897) new combination, major chelae (carpi and meri): A, left carpus, KGP- MH DH 065; B, right carpus, KGP-MH DH 060; C, left carpus, PCRB-DH 009, note serration on the lower margin (D); E, left articulated merus and carpus, SNM Z- 37549; F, left merus, PCRB-DH 014; G, right merus, KGP-MH DH 052; H, left merus, KGP-MH DH 010; I, right merus, PCRB-DH 016; J, left merus, PCRB-DH 015; K, right merus, SNM Z- 37542. All specimens belong to robust morphotype, except „ I “ being a slender morphotype. All specimens come from the Dúbravská hlavica locality (Vienna Basin, Slovakia), Middle Miocene. All specimens are to scale and were covered with ammonium chloride prior to photography.
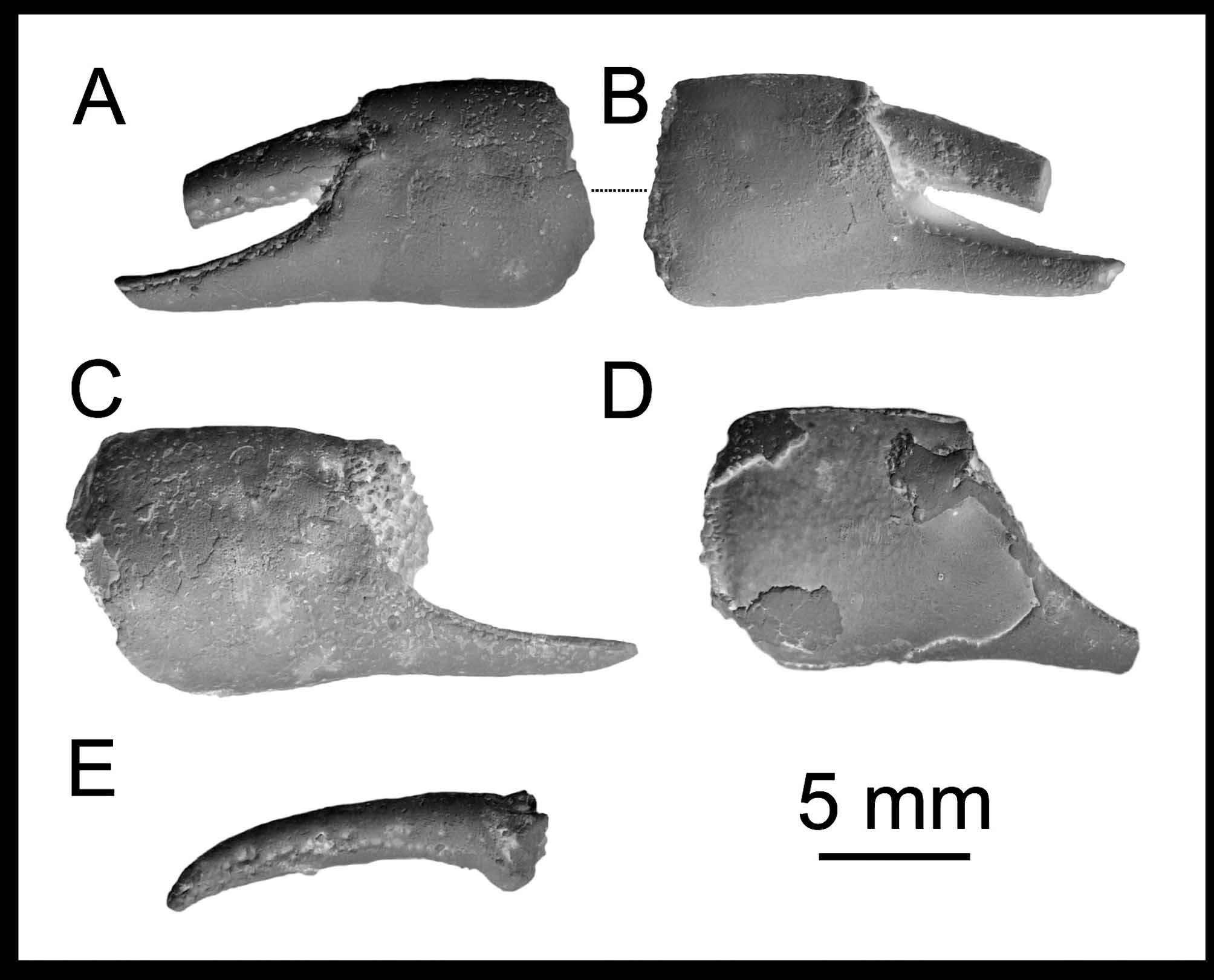
FIGURE 5. Neocallichirus brocchii (Lőrenthey, 1897) new combination, minor chelae (propodi and dactyli): A – B, left chela from outer and inner view, KGP-MH DH 067; C, right chela, PCRB-DH 013; D, fragmented right chela, PCRB-DH 011; E, left dactylus, PCRB-DH 012. All specimens come from the Dúbravská hlavica locality (Vienna Basin, Slovakia), Middle Miocene. All specimens are to scale and were covered with ammonium chloride prior to photography.

FIGURE 6. Neocallichirus brocchii (Lőrenthey, 1897) new combination, in situ preservation of the specimen KGP-MH DH- 0 58 (Dúbravská hlavica locality): A, view of the individual (slender morphotype) preserved in a presumed burrow structure; B, minor cheliped; C, major cheliped; D, view from below where both chelipeds can be recognized. Note also the partial preservation of thoracopods P 2 – P 4. All specimens are to scale and were covered with ammonium chloride prior to photography.
TABLE 1. Measurements of selected well preserved specimens of Neocallichirus brocchii (Lőrenthey, 1897) comb. nov. Handedness: L = left; R = right; measurements: L = max. length; H = max. height; ~ = approximate value;> = " more than " (in case the approximation is not possible due to the state of preservation); values are in mm.
| Specimen | morphotype | handedness | L | H | L | H | L H L |
|---|---|---|---|---|---|---|---|
| SNM Z-37532 | major robust | L | - | - | - | - | 14.0 14.4>9.4 |
| SNM Z-37533 SNM Z-37534 SNM Z-37535 | major robust major robust major robust | L L L | - - - | - - - | - - - | - - 15.7 | 10.8 18.3 - ~13>12 9.0 18.5 17.0>8.0 |
| SNM Z-37536 | major robust | L | - | - | - | - | 18.3 17.4 - |
| SNM Z-37537 | major robust | L | - | - | - | - | 13.0 13.2 - |
| SNM Z-37538 | major robust | L | - | - | - | - | - 15.0 - |
| SNM Z-37539 SNM Z-37540 | major robust major robust | L R | - - | - - | - - | - - | 11.4 11.4 - 14.7 15.0>8.0 |
| SNM Z-37541 | major robust | R | - | - | - | - | 14.4 14.0 - |
| SNM Z-37542 | major robust | R | - | - | - | - | 17.6 14.2 - |
| SNM Z-37543 SNM Z-37544 SNM Z-37545 | major robust major robust major robust | R R R | - - - | - - - | - - - | - - - | 13.2 12.2 - 17.0 17.2 - 18.0 17.0 - |
| SNM Z-37546 | major robust | L | - | - | - | - | - - 12.6 |
| SNM Z-37547 | major robust | L | - | - | 11.3 | 14.1 | - - - |
| SNM |
Slovak National Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Neocallichirus Sakai, 1988
| Hyžný, Matúš & Hudáčková, Natália 2012 |
Callianassa
| Hyzny 2011: 41 |
| Hyzny 2011: 167 |
Callianassa
| Muller 1984: 51 |
Callianassa brocchii.
| Schweitzer 2010: 34 |
| Muller 1979: 274 |
Callianassa
| Glaessner 1929: 77 |
Calianassa
| Lorenthey 1929: 69 |
Calianassa Brocchii Lőrenthey, 1897 : 161
| Lorenthey 1929: 64 |
| Lorenthey 1898: 106 |
| Lorenthey 1898: 132 |
| Lorenthey 1898: 104 |
| Lorenthey 1897: 161 |
1 (by plazi, 2016-04-12 07:35:44)
2 (by ImsDioSync, 2016-12-19 15:08:03)
3 (by ImsDioSync, 2016-12-19 15:09:20)
4 (by ExternalLinkService, 2019-09-26 19:48:34)
5 (by ExternalLinkService, 2021-10-29 01:38:07)
6 (by ExternalLinkService, 2021-10-29 04:00:38)
7 (by ExternalLinkService, 2021-10-29 14:53:30)
8 (by plazi, 2023-10-26 04:10:47)